|
Revista de Ciencias Ambientales (Trop J Environ Sci) |
NOTA TÉCNICA
Tolerancia ante inundaciones por parte de semillas y plantas de tres especies forestales nativas de América del Sur
Floods Tolerance of Seeds and Seedlings of Three Native Forest Species of South America
Victoria R. Kubota1, Derlis A. Britez2, Gabriel N. Duete3, Manuel M. Enciso G.4, María Lis García5, Sara B. Guillén6
[Recibido: 23 de setiembre 2019, Aceptado: 29 de enero 2020, Corregido: 22 de febrero 2020, Publicado: 1 de julio 2020]
Resumen
[Introducción]: En las últimas décadas, fueron registradas intensas precipitaciones e inundaciones en varios países de América del Sur, afectando los bosques ribereños y de galería. Sin embargo, los conocimientos sobre la tolerancia de las especies forestales a inundaciones aún son escasos. [Objetivo]: El objetivo de esta investigación fue evaluar el comportamiento de semillas y plantas de Luehea divaricata Mart., Parapiptadenia rigida (Benth.) Brenan y Guibourtia hymenaeifolia (Moric.) J. Léonard a las inundaciones en condiciones ex situ. [Metodología]: Las semillas de las especies seleccionadas fueron sometidas a inmersión en agua durante 7 distintos periodos de tiempo. Luego, las semillas fueron sembradas y se evaluó su porcentaje de germinación y la sobrevivencia de las plantas a los 30 días. [Resultados]: La tasa de germinación de L. divaricata se redujo de 45.9 % a 23.1 % durante el ensayo. Una quinta parte de las semillas de esta especie mantuvo el poder germinativo aún con 30 días de inmersión. Sin embargo, P. rigida y G. hymenaeifolia perdieron drásticamente el poder germinativo y de sobrevivencia cuando el periodo de inmersión superó los 5 días. [Conclusiones]. La respuesta de las semillas a inmersiones varió entre las especies, L. divaricata fue la que tuvo mayor tolerancia a condiciones de inundación.
Palabras clave: Ensayo de inmersión ex situ; Guibourtia hymenaeifolia (Moric.) J. Léonard; Luehea divaricata Mart.; Parapiptadenia rigida (Benth.) Brenan; sobrevivencia de plantas.
Abstract
[Introduction]: In the last decades, several countries of South America registered intense rainfalls and floods affecting riparian and gallery forests. Yet knowledge of the flooding tolerance of forest species is very limited. [Objective]: The aim of this study was to evaluate seeds and seedlings behavior of Luehea divaricata Mart., Parapiptadenia rigida (Benth.) Brenan, and Guibourtia hymenaeifolia (Moric.) J. Léonard subjected to ex situ flooding regimes. [Methods]: Seeds of the selected species were immersed in water over 7 different periods. After immersion seeds were sown and their germination percentage and seedlings survival were evaluated for 30 days. [Results]: L. divaricata reduced its germination rate during the experiment from 45.9 % to 23.1 %. One fifth of seeds of this species kept the germination capability even after 30 days of the immersion treatment. A visible loss of the germination capability and seedling survival was observed in P. rigida and G. hymenaeifolia when the immersion period exceeded 5 days. [Conclusions]: The response of seeds to immersions varied between species, being L. divaricata the species with better tolerance to flooding condition.
Keywords: ex situ immersion experiment; Guibourtia hymenaeifolia (Moric.) J. Léonard; Luehea divaricata Mart.; Parapiptadenia rigida (Benth.) Brenan; seedling survival
En las últimas décadas se registró un aumento de precipitaciones en varios países de América del Sur, como en el sureste de Brasil, Paraguay, Uruguay, partes de Argentina y de Bolivia (Céspedes y Ríos, 1985; Kaenzig y Piguet, 2014). Estas variantes en la precipitación pueden aumentar la intensidad o la frecuencia de las inundaciones (Arnell y Gosling, 2016). Las proyecciones realizadas estiman alteraciones mayores en el régimen de precipitación a causa del cambio climático, lo cual sugiere el posible incremento de inundaciones en varias regiones del mundo (Arnell y Gosling, 2016; Bidegain et al., 2012; Hirabayashi et al., 2013). Los cauces hídricos y sus zonas de influencia son las más afectadas por estos eventos naturales (Chamorro, 2010).
Los bosques ribereños y en galería cumplen importantes funciones en el régimen hídrico, además de ofrecer servicios ecosistémicos de relevancia para la ciudadanía (Vidal, 2010). Es de esperar que las especies forestales que componen estos bosques toleren las condiciones de alto contenido de humedad. Aun así, los ambientes propicios para la generación de patógenos y la privación de oxígeno, sumados a otros impactos negativos causados por las inundaciones prolongadas, pueden afectar la vegetación.
El efecto de las inundaciones puede incidir en la etapa de germinación y establecimiento de las plantas. El conocimiento sobre la tolerancia de las semillas y plantas de las especies forestales ante las inundaciones es fundamental para estimar la capacidad de subsistencia y recuperación de los ecosistemas propensos a eventos de esta índole; esta información ayudaría a mejorar el manejo adaptativo.
Estudios realizados a nivel regional indican que la tolerancia de las semillas y plantas de las especies forestales a inundaciones es muy variable (Parolin, 2002; Scarano et al., 1997; Scarano, 1998). Pese a la existencia de ecosistemas forestales propensos a inundaciones, en la región aún no se conoce suficientemente el comportamiento de las especies nativas ante eventos de esta índole. En este contexto, surge la necesidad de estudiar la capacidad de subsistencia y recuperación de los ecosistemas ribereños y en galerías.
Luehea divaricata Mart. es una especie selectivamente higrófita de la familia Malvaceae (anteriormente clasificada como Tiliaceae) que abunda en los sitios húmedos y bajos; se encuentra en los bosques ribereños y de galería de Paraguay, Brasil, Argentina y Uruguay. Su fruto es una cápsula leñosa dehiscente que contiene numerosas semillas aladas de 5-7 mm (López et al., 2002; Lorenzi, 2014a).
Parapiptadenia rigida (Benth.) Brenan es una especie de la familia Fabaceae que prefiere el suelo arcilloso y crece tanto en suelo húmedos como secos de Paraguay, Brasil, Bolivia, Argentina y Uruguay. Su fruto es una vaina dehiscente que contiene 5-15 semillas planas de 8-10 mm, rodeadas por un ala angosta (López et al., 2002; Lorenzi, 2014a; Missouri Botanical Garden, 2020; Peña-Chocarro et al., 2006).
Guibourtia hymenaeifolia (Moric.) J. Léonard. es una especie de la familia Fabaceae que crece en Bolivia, Brasil y Paraguay; habita suelos arenosos en sitios donde factores edáficos impiden el desarrollo del bosque alto; abundan en las cercanías de arroyos. Su fruto es una vaina dehiscente que contiene una semilla dura de 1 cm aproximadamente (López et al., 2002; Lorenzi, 2014b; Missouri Botanical Garden, 2020).
El estudio tuvo como objetivo evaluar el comportamiento germinativo de las semillas y la supervivencia de las plantas de L. divaricata, P. rigida y G. hymenaeifolia ante las inundaciones en condiciones ex situ.
El estudio fue realizado en el vivero forestal de la Facultad de Ciencias Agrarias de la Universidad Nacional de Asunción, ubicado en la ciudad de San Lorenzo, Paraguay, durante el periodo de septiembre a noviembre de 2016.
Se seleccionaron tres especies forestales nativas que habitan en zonas de influencia de los cauces hídricos de algunos países de América del Sur: L. divaricata, P. rigida y G. hymenaeifolia. Se emplearon 2 800 semillas por especie que fueron colectadas de árboles semilleros que se encuentran en el Campo Experimental de la Facultad de Ciencias Agrarias, de la Universidad Nacional de Asunción (25° 19’ 36.24” S 57° 31’ 10.96” W). Los árboles semilleros de L. divaricata y G. hymenaeifolia fueron plantados entre los años 1989 y 1990, y forman parte del Arboretum de la mencionada institución. No se conoce la edad de los árboles de P. rigida, debido a que se trata de una regeneración natural instalada en el predio. Las semillas fueron colectadas en agosto y septiembre de 2016. Se eliminaron las impurezas y las semillas en buen estado fueron mezcladas para formar un único lote. Se inició el ensayo de inmersión justo después de estas labores.
Las tres especies fueron seleccionadas mediante los criterios: zona de distribución natural, disponibilidad de suficiente cantidad de semillas en el campo experimental y época de maduración de frutos.
Para simular el escenario de un campo inundado en condiciones ex situ, las semillas fueron sumergidas en agua por tiempo predefinido en envases de plástico (10 cm de altura y 8 cm de diámetro) que contenían suelo y agua de pozo artesiano hasta la mitad del recipiente. Un total de 400 semillas, divididas en 20 repeticiones de 20 semillas cada una, fueron sometidas a los distintos periodos de inmersión (ISTA, 2016); los lapsos fueron 0, 5, 10, 15, 20, 25 y 30 días. Los envases fueron ubicados bajo techo, a temperatura ambiente y sin iluminación artificial. La temperatura media mensual registrada para la zona fue de 21.4, 25.0 y 25.7 °C, y la humedad relativa de 55.3, 62.5 y 61.9 % respectivamente para septiembre, octubre y noviembre de 2016, periodo en el que se realizó el experimento. El agua de los envases fue reemplazada cada 2 días a fin de evitar la fermentación de las semillas.
Posterior a la inmersión en agua, durante los periodos predefinidos, se procedió al conteo de las semillas en las siguientes categorías: semillas en buen estado no germinadas, semillas germinadas y semillas con signos de pudrición. Las semillas que emitieron radícula o tallo en los envases de plástico fueron contabilizadas como germinadas en agua. Las semillas que se desintegraron en el agua fueron consideradas como podridas y fueron desechadas.
Las semillas en buen estado no germinadas y aquellas germinadas en agua fueron sembradas o trasplantadas en tablones de arena de 1 m de ancho para el estudio de germinación y sobrevivencia a campo abierto. Los tablones fueron cubiertos por media sombra de 50 % de luminosidad y se aplicaron 2 riegos de agua de pozo por día. El conteo de plantas germinadas y vivas se realizó a los 30 días después de la siembra o trasplante. A fin de diferenciar con el ensayo de germinación de semillas inmersas en agua, se denomina a éste como estudio de sobrevivencia de plantas en suelo.
Se determinó el porcentaje de germinación y de pudrición de semillas en envase de cada repetición de cada periodo de inmersión. Para comparar la media entre los periodos de inmersión y entre las especies, se aplicó el test de Kruskal-Wallis con un nivel de confianza del 95 %, y el test de Bonferroni cuando existe diferencia significativa entre las medias, con un nivel de confianza del 95 %. Se empleó el programa estadístico R versión 3.5.1 (R Core Team, 2018) para ambos análisis.
Además, se analizó la sobrevivencia de semillas y plantas inmersas en agua mediante la curva de supervivencia de Kaplan-Meier, empleando el programa estadístico R versión 3.5.1 (R Core Team, 2018). Para el efecto, se cuantificaron las semillas en buen estado no germinadas y las plantas germinadas en agua y vivas al final de cada periodo de inmersión. Algunas semillas de L. divaricata no fueron encontradas al final del periodo de inmersión, debido a que por su reducido tamaño se mezclaron con la arena, lo cual dificultó su visualización. Estas semillas no fueron incluidas en los conteos.
Posteriormente, se calculó el porcentaje de sobrevivencia de las plantas en suelo a los 30 días de la siembra o trasplante en tablón de cada repetición de cada periodo de inmersión. Para comparar la media entre los periodos de inmersión y entre las especies, se aplicó el test de Kruskal-Wallis con un nivel de confianza del 95 %. En caso de encontrarse diferencia significativa entre las medias, se aplicó el test de Bonferroni con un nivel de confianza del 95 %, con el programa estadístico R versión 3.5.1 (R Core Team, 2018).
3.1 Germinación y pudrición de semillas inmersas en agua
Hubo diferencia significativa entre la media del porcentaje de germinación de semillas inmersas en agua durante diferentes periodos, tanto de L. divaricata como de P. rigida (p < 0.05; Cuadro 1, en horizontal). Asimismo, hubo diferencia significativa entre la media de porcentaje de germinación de las especies (p < 0.05; Cuadro 1, en vertical).
Cuadro 1. Media de porcentaje de semillas germinadas en agua, comparación de media de porcentaje de germinación entre diferentes periodos de inmersión (valor p < 0.05) y entre las especies (valor p < 0.05)
Table 1. Means of percentage of seeds germinated in water, comparison of means of germination percentage between different periods of immersion (p value <0.05) and between species (p value <0.05)
|
Media de porcentaje de semillas germinadas en agua |
||||||||||||
|
|
5 d |
10 d |
15 d |
20 d |
25 d |
30 d |
||||||
|
L. divaricata |
0.0 |
± 0.0 |
5.3 |
± 0.9 |
10.8 |
± 1.4 |
10.8 |
± 1.4 |
14.0 |
± 1.9 |
12.8 |
± 1.7 |
|
P. rigida |
100.0 |
± 0.9 |
96.3 |
± 0.9 |
96.5 |
± 0.7 |
96.5 |
± 0.8 |
96.5 |
± 0.9 |
95.5 |
± 0.8 |
|
G. hymenaeifolia |
0.0 |
± 0.0 |
0.0 |
± 0.0 |
0.0 |
± 0.0 |
0.0 |
± 0.0 |
0.0 |
± 0.0 |
0.0 |
± 0.0 |
|
Comparación de media de porcentaje de germinación entre diferentes periodos de inmersión * |
||||||||||||
|
|
5 d |
10 d |
15 d |
20 d |
25 d |
30 d |
||||||
|
L. divaricata |
a |
b |
bc |
bc |
c |
c |
||||||
|
P. rigida |
a |
b |
ab |
b |
b |
b |
||||||
|
G. hymenaeifolia |
- |
- |
- |
- |
- |
- |
||||||
|
Comparación de media de porcentaje de germinación entre especies ** |
||||||||||||
|
|
5 d |
10 d |
15 d |
20 d |
25 d |
30 d |
||||||
|
L. divaricata |
a |
a |
a |
a |
a |
a |
||||||
|
P.rigida |
b |
b |
b |
b |
b |
b |
||||||
|
G. hymenaeifolia |
a |
c |
c |
c |
c |
c |
||||||
Notas: d= días; * Resultados del test de Bonferroni en horizontal; ** Resultados del test de Bonferroni en vertical.
Por otra parte, se observó diferencia significativa entre la media del porcentaje de semillas podridas de los periodos en P. rigida y G. hymenaeifolia (p < 0.05; Cuadro 2, en horizontal). Así también, se encontró diferencia significativa entre la media de porcentaje de pudrición de semillas de las especies en cada periodo de inmersión (p < 0.05; Cuadro 2, en vertical).
Cuadro 2. Media de porcentaje de semillas que presentaron signo de pudrición, comparación de media de porcentaje de semillas con signos de pudrición entre diferentes periodos de inmersión (valor p < 0.05) y entre las especies (valor p < 0.05)
Table 2. Means of percentage of seeds with rot signs, comparison of means of percentage of seeds with rot signs between different periods of immersion (p <0.05) and between species (p <0.05)
|
Media de porcentaje de semillas podridas en agua |
||||||||||||
|
|
5 d |
10 d |
15 d |
20 d |
25 d |
30 d |
||||||
|
L. divaricata |
0.0 |
± 0.0 |
0.0 |
± 0.0 |
0.0 |
± 0.0 |
0.0 |
± 0.0 |
0.0 |
± 0.0 |
0.0 |
± 0.0 |
|
P. rigida |
0.0 |
± 0.0 |
3.8 |
± 0.9 |
3.5 |
± 0.7 |
3.5 |
± 0.8 |
3.5 |
± 0.9 |
4.5 |
± 0.8 |
|
G. hymenaeifolia |
0.0 |
± 0.0 |
7.8 |
± 1.0 |
12.8 |
± 1.5 |
30.5 |
± 1.7 |
46.0 |
± 2.6 |
63.5 |
± 2.6 |
|
Comparación de media de porcentaje de semillas podridas entre diferentes periodos de inmersión* |
||||||||||||
|
|
5 d |
10 d |
15 d |
20 d |
25 d |
30 d |
||||||
|
L. divaricata |
- |
- |
- |
- |
- |
- |
||||||
|
P. rigida |
a |
b |
ab |
b |
b |
b |
||||||
|
G. hymenaeifolia |
a |
b |
b |
c |
d |
e |
||||||
|
Comparación de media de porcentaje de semillas podridas entre especies ** |
||||||||||||
|
|
5 d |
10 d |
15 d |
20 d |
25 d |
30 d |
||||||
|
L. divaricata |
- |
a |
a |
a |
a |
a |
||||||
|
P. rigida |
- |
b |
b |
b |
b |
b |
||||||
|
G. hymenaeifolia |
- |
c |
c |
c |
c |
c |
||||||
Notas: d= días; * Resultados del test de Bonferroni en horizontal; ** Resultados del test de Bonferroni en vertical.
Las semillas de L. divaricata empezaron a germinar en el envase a los 5 días de inmersión en agua (Cuadro 1). Los porcentajes de germinación en el envase fueron inferiores al 15 % para todos los lapsos de inmersión. Ninguna semilla inmersa presentó señales de pudrición, lo cual demostró su alta tolerancia al agua (Cuadro 2 y Figura 1). Una vez germinadas, las plantas presentaron visible alargamiento del tallo sin perder la viabilidad. Todas las plantas germinadas sobrevivieron en agua hasta el momento del trasplante al tablón (Figura 1).
La mayoría de las semillas de P. rigida empezó a germinar antes de los 5 días de inmersión en agua (Cuadro 1). Fueron pocas las semillas de P. rigida que se pudrieron en el envase sin germinar (3.0 % en total) (Cuadro 2). El mayor porcentaje de pudrición de semillas de P. rigida fue registrada en aquellas con 30 días de inmersión (4.5 %). Sin embargo, solo las plantas inmersas por 5 días estuvieron en buen estado en el momento del trasplante al tablón (Figura 1). La mayoría de los individuos germinados que permanecieron inmersos en agua por más días presentaron debilitamiento o signos de pudrición al momento de ser trasplantados al tablón.
Las semillas de G. hymenaeifolia presentaron aumento de volumen y ruptura de tegumento, pero ninguna germinó en el agua (Cuadro 1). El agua de los envases con las semillas de esta especie presentó un color oscuro y un aspecto gelatinoso. El número de semillas de G. hymenaeifolia con signos de pudrición fue aumentando gradualmente a medida que se prolongaba el periodo de inmersión, y llegó a 63.5 % después de los 30 días de inmersión (Cuadro 2, Figura 1).
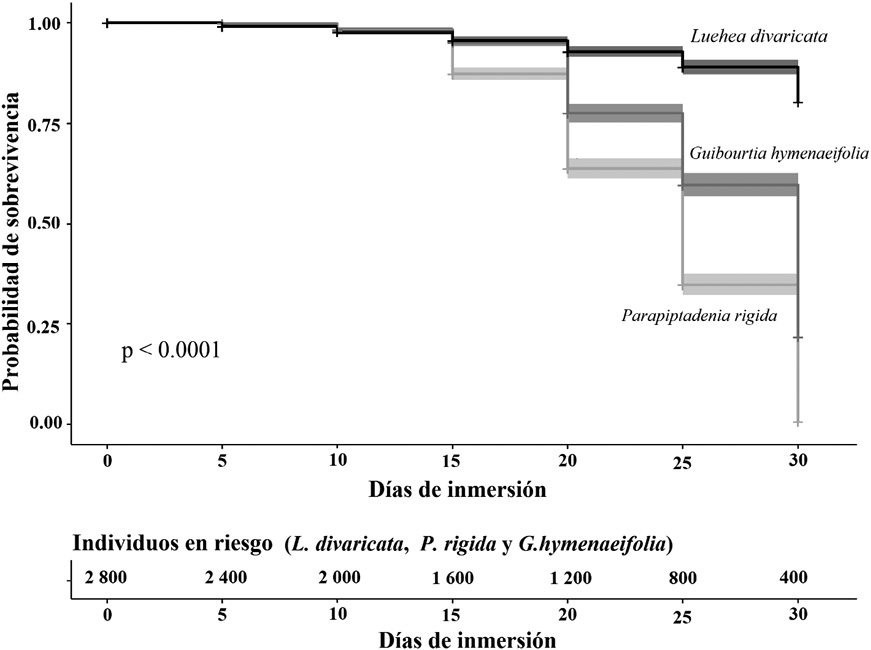
Figura 1. Curva de sobrevivencia de las semillas y plantas germinadas en agua.
Figure 1. Survival curve of seeds and plants germinated in water.
3.2 Sobrevivencia de plantas en suelo
Hubo diferencia significativa entre la media del porcentaje de sobrevivencia de las plantas a los 30 días después de la siembra o trasplante (en caso de aquellas plantas germinadas en el envase) de los periodos de inmersión de cada especie (p < 0.05; Cuadro 3, en horizontal). Asimismo, se observó diferencia significativa entre la media de porcentaje de sobrevivencia a los 30 días después de la siembra o trasplante de las especies en cada periodo de inmersión (p < 0.05; Cuadro 3, en vertical).
El porcentaje de sobrevivencia de las plantas a los 30 días de la siembra o trasplante de L. divaricata presentó un descenso gradual con el alargamiento del periodo de inmersión previo. Por su parte, el porcentaje de sobrevivencia en tablón de G. hymenaeifolia y P. rigida se redujo visiblemente cuando el periodo de inmersión previa en agua superó los 10 días. Se observaron diferencias significativas entre G. hymenaeifolia y P. rigida; no obstante, demostraron tendencias similares en el porcentaje de plantas que sobrevivieron hasta los 30 días de la siembra o trasplante y una tendencia diferente a L. divaricata (Cuadro 3).
Cuadro 3. Media de porcentaje de sobrevivencia de plantas a los 30 días de la siembra en tablón, comparación de media de porcentaje de sobrevivencia de plantas entre los periodos de inmersión previa (valor p < 0.05) y entre las especies (valor p < 0.05)
Table 3. Means of plants survival percentage 30 days after sowing in ground, comparison of means of plants survival percentage between previous immersion periods (p value <0.05) and species (p value <0.05)
|
Media de porcentaje de sobrevivencia de plantas |
||||||||||||||
|
|
0 d |
5 d |
10 d |
15 d |
20 d |
25 d |
30 d |
|||||||
|
L. divaricata |
42.3 |
± 13.6 |
45.9 |
± 17.2 |
44.0 |
± 14.3 |
34.0 |
± 8.9 |
34.0 |
± 11.8 |
27.0 |
± 18.4 |
23.1 |
± 14.7 |
|
P. rigida |
79.8 |
± 3.5 |
73.8 |
± 1.9 |
41.5 |
± 2.4 |
8.8 |
± 1.2 |
8.8 |
± 0.6 |
4.3 |
± 27.8 |
0.0 |
± 0 |
|
G. hymenaeifolia |
88.5 |
± 9.6 |
80.5 |
± 10.5 |
29.0 |
± 10.1 |
4.8 |
± 15.7 |
4.8 |
± 5.4 |
0.8 |
± 2.4 |
0.0 |
± 0 |
|
Comparación de media de porcentaje de plantas vivas entre diferentes periodos de inmersión previa* |
||||||||||||||
|
|
0 d |
5 d |
10 d |
15 d |
20 d |
25 d |
30 d |
|||||||
|
L. divaricata |
a |
a |
ab |
c |
abd |
acd |
cd |
|||||||
|
P. rigida |
a |
a |
b |
c |
d |
d |
e |
|||||||
|
G. hymenaeifolia |
a |
a |
b |
c |
cd |
de |
e |
|||||||
|
Comparación de media de porcentaje de plantas vivas entre especies ** |
||||||||||||||
|
|
0 d |
5 d |
10 d |
15 d |
20 d |
25 d |
30 d |
|||||||
|
L. divaricata |
a |
a |
a |
a |
a |
a |
a |
|||||||
|
P. rigida |
b |
b |
a |
b |
b |
b |
b |
|||||||
|
G. hymenaeifolia |
b |
b |
b |
c |
b |
b |
b |
|||||||
Notas: d= días; * Resultados del test de Bonferroni en horizontal; ** Resultados del test de Bonferroni en vertical.
La respuesta de las semillas y plantas de L. divaricata registrada en este estudio es similar al comportamiento de las semillas de Carapa guianensis reportado por Scarano et al. (2003); en ensayos de inundación ex situ, las plantas de C. guianensis que germinaron en agua presentaron crecimiento longitudinal, sin perder la viabilidad, a pesar del retraso en el desarrollo de las hojas verdaderas. Cabe destacar que C. guianensis es una especie arbórea de los bosques lluviosos de amazonía, capaz de crecer tanto en bosques bien drenados como en bosques propensos a inundaciones. Parolin (2002) también reportó que varias especies amazónicas desarrollan tallos alargados como estrategias de adaptación a las inundaciones.
El comportamiento de las semillas de L. divaricata y C. guianensis sugiere que las especies forestales que crecen en bosques propensos a inundaciones pueden tener la habilidad de germinar estando inmersas en agua. El efecto de la inmersión puede manifestarse en la morfología de las plantas y el retraso de su desarrollo, pero sin afectar la sobrevivencia, propiedades que hacen a estas especies tolerantes a condiciones de inundaciones.
El porcentaje de germinación registrado para L. divaricata fue bajo en comparación con las demás especies, 45.9 % ± 17.2 fue el máximo valor alcanzado con 5 días de inmersión. En estudios realizados con la misma especie por Dutra et al. (2016), y Borges et al., (2007), fueron reportados porcentajes de germinación de 42 % y 51.9 %, respectivamente. Estos resultados sugieren que L. divaricata podría ser una especie con baja capacidad de germinación.
A pesar de que G. hymenaeifolia y P. rigida presentaron tendencias similares en el porcentaje de plantas germinadas que sobrevivieron hasta los 30 días de la siembra o trasplante, ambas especies demostraron comportamientos diferentes durante el ensayo de inmersión en agua. Se observó un elevado porcentaje de germinación de P. rigida a los 5 días de inmersión. Esta tendencia coincide con lo reportando por Lorenzzi (2014a), quien indicó un elevado porcentaje de germinación de la misma especie a los 5 a 10 días después de la siembra. Mientras que las semillas de G. hymenaeifolia indicaron poca tolerancia a la inmersión en agua, con nula germinación y elevado porcentaje de semillas con signos de pudrición en esa condición. En condiciones normales, G. hymenaeifolia presenta bajo porcentaje de germinación, que tiene lugar aproximadamente a los 35 días después de la siembra (Lorenzzi, 2014b).
Los resultados sugieren que L. divaricata tiene alta tolerancia a las inundaciones, debido a que sus semillas tardan en perder su poder germinativo y las plantas pueden sobrevivir a pesar de permanecer inmersas en agua. Por otra parte, las inundaciones durante menos de 5 días pueden favorecer la germinación de P. rigida. Sin embargo, la sobrevivencia de las plantas de esta especie se reduce rápidamente, si permanecen inmersas en agua por más de 5 días. G. hymenaeifolia es una especie que no tolera las inundaciones, pues la presencia del agua propicia la pudrición de sus semillas.
Las diferencias de tendencias de germinación y sobrevivencia observadas pueden deberse a que L. divaricata es una especie propia de los bosques ribereños (López et al., 2002), y que sus semillas y plantas son más tolerantes a inmersiones prolongadas. Mientras que P. rigida y G. hymenaeifolia, aparte de crecer en bosques ribereños y en galería, se producen también en suelos secos (López et al., 2002), en donde la posibilidad de que las semillas estén en contacto con el agua durante un periodo prolongado es menor.
La tolerancia de las semillas y plantas ante las inundaciones varía entre las especies estudiadas. L. divaricata tiene la mayor resistencia a inmersión en agua por periodo prolongado. Sus semillas tardan en perder el poder germinativo y las plantas germinadas en agua tienen la capacidad de sobrevivir en condiciones de anegamiento. P. rigida y G. hymenaeifolia no resisten ante las inundaciones por periodos prolongados.
Al Fondo de Investigación de la Dirección de Investigación Científica y Tecnológica del Rectorado de la Universidad Nacional de Asunción, por financiar este estudio. A las estudiantes universitarias Rosa Colman y Paola Lezcano y a los funcionarios del Vivero Forestal de la Facultad de Ciencias Agrarias de la Universidad Nacional de Asunción, por apoyar la ejecución de los ensayos. Finalmente, a las personas revisoras anónimas, por sus oportunos comentarios al escrito.
7.Ética y conflicto de intereses
Las personas autoras declaran que han cumplido totalmente con todos los requisitos éticos y legales pertinentes, tanto durante el estudio como en la producción del manuscrito; que no hay conflictos de intereses de ningún tipo; que todas las fuentes financieras se mencionan completa y claramente en la sección de agradecimientos; y que están totalmente de acuerdo con la versión final editada del artículo.
Arnell, W. N. y Gosling, N. S. (2016). The impacts of climate change on river flood risk at the global scale. Climatic Change, 134, 387-401. https://doi.org/10.1007/s10584-014-1084-5
Bidegain, M., Coronel, G., Ríos, N. y Santos, D. (2012). Escenarios climáticos futuros para Paraguay. Meteorológica, 37(2), 47-55.
Borges, K. C. F., Santana, D. G., Ranal, M., Dorneles, M. C. y Carvalho, M. P. (2007). Germinação de sementes e emergência de plántulas de Luehea divaricata Mart. Revista Brasileira de Biociências, 5(2), 1008-1010.
Céspedes, R. L. y Ríos, L. S. (1985). Análisis del impacto de las inundaciones en el Paraguay. http://cidbimena.desastres.hn/docum/crid/Marzo2004/pdf/spa/doc1018/doc1018.htm
Chamorro Vega, L. F. (2010). Análisis estadístico de la variabilidad climática y sus impactos en Paraguay. Universidad Nacional de Asunción. http://sdi.cnc.una.py/catbib/documentos/442.pdf
Dutra, A. F., Machado Araujo, M., Guarienti Rorato, D. y Mieth, P. (2016). Germinação de sementes e emergência de plântulas de Luehea divaricata Mart. et. Zucc. em diferentes substratos. Ciência Florestal, 26(2), 411-418. https://doi.org/10.5902/1980509822744
Hirabayashi, Y., Mahendran, R., Koirala, S., Konoshima, L., Yamazaki, D., Watanabe, S., Kim, H. y Kanae, S. (2013). Global flood risk under climate change. Nature Climate Change, 3(9), 816-821. https://doi.org/10.1038/nclimate1911
International Seed Testing Association (ISTA). (2016). International Rules for Seed Testing. https://www.seedtest.org
Kaenzig, R. y Piguet, E. (2014). Migration and climate change in Latin America and the Caribbean. En E. Piguet y F. Laczko (Eds.), People on the move in a changing climate (pp. 155-176). https://doi.org/10.1007/978-94-007-6985-4_7
López, J. A., Little, E. L., Ritz, G. F., Rombold, J. y Hahn, W. J. (2002). Árboles comunes del Paraguray: Ñande yvyra mata kuera. FCA-UNA y Cuerpo de Paz.
Lorenzi, H. (2014a). Árvores brasileiras: Manual de identificação e cultivo de plantas arbóreas nativas do Brasil (Vol. 1). Plantarum.
Lorenzi, H. (2014b). Árvores brasileiras: Manual de identificação e cultivo de plantas arbóreas nativas do Brasil (Vol. 2). Plantarum.
Missouri Botanical Garden. (2020). Trópicos. http://www.tropicos.org/nameSearch
Parolin, P. (2002). Submergence tolerance vs. escape from submergence: two strategies of seedling establishment in Amazonian floodplains. Tropical Ecology, 48(2), 177-186. https://doi.org/10.1016/S0098-8472(02)00036-9
Peña-Chocarro, M. C., De Egea Juvenil, J., Vera, M., Maturo, H. y Knapp, S. (2006). Guía de árboles y arbustos del Chaco húmedo. The Natural History Museum, Guyra Paraguay, Fundación Moisés Bertoni y Fundación Hábitat y Desarrollo.
R Core Team. (2018). R: A language and environment for statistical computing. R Foundation for Statistical. Computing, Vienna, Austria. URL https://www.R-project.org
Scarano, F. R., Ribeiro, K. T., Moraes, L. F. D. y Lima, H. C. (1997). Plant establishment on flooded and unflooded patches of a freshwater swamp forest in southeastern Brazil. Journal of Tropical ecology, 13(6), 793-803. https://doi.org/10.1017/S0266467400011007
Scarano, F. R. (1998). A comparison of dispersal, germination and establishment of woody plants subjected to distinct flooding regimes in Brazilian flood-prone forests and estuarine vegetation. En F. R. Scarano y A. C. Franco (Eds.), Ecophysioogical strategies of xerophytic and amphibious plants in the neotropics (pp. 177-193). Series Oecologia Brasilienses. http://www.oecologiaaustralis.org/ojs/index.php/oa/article/view/285/355. https://doi.org/10.4257/oeco.1998.0401.09
Scarano, F. R., Pereira, T. S. y Rôças, G. (2003). Seed germination during flotation and seedling growth of Carapa guianensis, a tree from flood-prone forests of the Amazon. Plant Ecology, 168(2), 291-296. http://link.springer.com/article/10.1023/A:1024486715690#page-1. https://doi.org/10.1023/A:1024486715690
Vidal, V. S. (2010). Eje estratégico desarrollo forestal sostenible y servicios ambientales. FAO. http://www.silvapar.com/politica-forestal/1.%20Forestal%20general/2.2.%20MAG%20-%20MARCO%20EJE%20ESTRATEGICO%20Vidal%20Agosto%2010.pdf
1 Ingeniera forestal, académica de la Facultad de Ciencias Agrarias, Universidad Nacional de Asunción, Paraguay; kubotapy@agr.una.py; https://orcid.org/0000-0001-7080-7406
2 Estudiante de la carrera de Ingeniería Forestal, Facultad de Ciencias Agrarias, Universidad Nacional de Asunción, Paraguay; derlibrite654@gmail.com; https://orcid.org/0000-0002-9403-9998
3 Estudiante de la carrera de Ingeniería Forestal, Facultad de Ciencias Agrarias, Universidad Nacional de Asunción, Paraguay; gabroduete@hotmail.com; https://orcid.org/0000-0002-8172-4507
4 Ingeniero forestal, académico de la Facultad de Ciencias Agrarias, Universidad Nacional de Asunción, Paraguay; manuelenci@yahoo.com; https://orcid.org/0000-0003-0439-5655
5 Ingeniera forestal, académica de la Facultad de Ciencias Agrarias, Universidad Nacional de Asunción, Paraguay; marialisgarcia@gmail.com; https://orcid.org/0000-0003-2811-6585
6 Estudiante de la carrera de Ingeniería Forestal, Facultad de Ciencias Agrarias, Universidad Nacional de Asunción, Paraguay; sariikusa@gmail.com; https://orcid.org/0000-0002-1763-6852



