
Revista de Ciencias Ambientales (Trop J Environ Sci). (Junio, 2015). EISSN: 2215-3896. Vol 49(1): 1-15.
DOI: http://dx.doi.org/10.15359/rca.49-1.1
URL: www.revistas.una.ac.cr/ambientales
EMAIL: revista.ambientales@una.cr

Variación del contenido foliar de nutrimentos de Gmelina arborea en los cantones de Osa, Golfito y Corredores, Costa Rica
Variation of Foliar Content of Nutriments of Gmelina arborea in the Counties of Osa, Golfito and Corredores, Costa Rica
María José Avellán-Zumbado a, Rafael Murillo-Cruz b, Alfredo Alvarado-Hernández c,
Carlos Ávila-Arias d
a Licenciada en Ciencias Forestales, investigadora de la Universidad Nacional de Costa Rica (UNA), maria.avellan.zumbado@una.cr;
b Máster en Ciencias Agrícolas y Recursos Naturales con énfasis en Suelos, investigador y académico, Instituto de Investigación y Servicios Forestales, UNA, murillorafael5454@yahoo.com;
c Ingeniero Agrónomo, profesor emérito de la Universidad de Costa Rica, alfredo.alvarado@ucr.ac.cr;
d Investigador y académico; Máster en Gestión de Recursos Naturales y Tecnologías de Producción, Instituto de Investigación y Servicios Forestales, UNA, carlosenriquea79@gmail.com
[Recibido: 23 de agosto 2015; Aceptado: 16 de octubre 2015; Corregido: 2 de noviembre 2015 Publicado: 29 de febrero 2016]
Resumen
Se evaluó la variación de la concentración de los nutrimentos foliares en 12 plantaciones de G. arborea (melina) con edades de 1-12 años, de los cantones de Osa, Golfito y Corredores en Costa Rica. Se realizó un muestreo foliar de la copa superior externa, superior interna, media externa y copa media interna, en la época lluviosa y seca. Se seleccionaron 45 árboles dominantes (2 árboles por época por edad). A cada muestra se le realizó un análisis químico completo y aluminio. Se obtuvieron variaciones en las concentraciones foliares debidas a la época de muestreo, y resultó mayor el N (2,99%), K (1,14%), Fe (91 mg kg-1) y Al (53 mg kg-1) en la época lluviosa, y Ca (1,42%) y Mn (95 mg kg-1) en la seca. Las concentraciones no variaron según la posición de las hojas en la copa, a excepción del Ca, que presentó mayores concentraciones en la copa media interna (1,40%) respecto a la superior externa (1,02%). La edad causó variaciones en Ca, K, B y Fe. Se estableció un protocolo de muestreo foliar y se determinaron los rangos de concentración foliar de macro y micronutrimentos para la especie en la Zona Sur de Costa Rica.
Palabras claves: Análisis foliar, macronutrimentos, micronutrimentos, muestreo foliar, rangos de concentración foliar.
Abstract
We assessed the variation of foliar nutriments concentration in 12 plantations of G. arborea (melina) with ages between 1-12 years old, of the Cantons of Osa, Golfito and Corredores in Costa Rica. A foliar sampling of outer upper-top, inner top, outer half and inner half-top was carried out in the rainy and dry seasons. Forty five dominant trees (2 trees by season by age) were selected. A full chemical analysis and aluminum was performed in each sample. Variations in foliar concentrations due to the time of sampling were obtained with a higher N (2.99%), K (1.14%), Fe (91 mgkg- 1) and Al (53 mgkg- 1) in the rainy season, and Ca (1.42%) and Mn (95 mgkg- 1) in the dry season. Concentrations did not vary according to the position of the leaves in the top, with the exception of the Ca, which presented higher concentrations in the inner half-top (1.40%) with respect to the outer upper-top (1.02%). The age caused variations in Ca, K, B and Fe. A foliar sampling protocol was established and determined foliar concentration ranges of macro and micronutrients for the species in the Costa Rican Southern area.
Keywords: Foliar analysis, foliar concentration ranges, leaf sampling, macronutrients, micronutrients.
- Introducción
La concentración de nutrimentos en tejidos vegetales se determina, principalmente, en las hojas por medio de su digestión total (Binkley, 1993; Munson y Nelson, 1973). Esta herramienta proporciona información sobre el estado nutricional de las plantas (Correndo y García, 2012), permite diagnosticar y predecir posibles trastornos de nutrimentos (Evans, 1979; Smith y Loneragan, 1997), y evaluar la efectividad de los programas de fertilizaciones (Aldrich, 1973; Brockley, 2001). Para obtener buenos resultados del análisis, se deben estandarizar las técnicas de muestreo e identificar los factores que provocan variación en las concentraciones foliares de nutrimentos, entre ellos la edad, la especie, el tejido y el nutrimento (Correndo y García, 2012). El uso de esta herramienta es poco común en plantaciones de especies tropicales de madera dura, debido a la variedad de especies, los largos turnos de corta, y su adaptación a gran variedad de suelos y climas (en particular con diferentes niveles de estrés hídrico), lo que limita la definición de los contenidos óptimos de nutrimentos de estas mismas (Brunck, 1987).
La Gmelina arborea (melina) es una especie nativa de países del sureste de Asia, introducida en Costa Rica en el año 1966 en la zona Atlántica por la empresa Celulosa de Turrialba (Lega, citado por Rojas y Murillo, 2004). Esta especie ha presentado una buena adaptación, un rápido crecimiento y diversidad de usos (Arce y Barrantes, 2006), lo cual la posiciona como una de las más plantadas en Costa Rica.
Existen algunos estudios de concentraciones foliares de nutrimentos en melina (Chijioke, 1980; Drechsel y Zech, 1991; Evans, 1979; González, 1996; Sampaio y Huber, 1999; Stuhrmann, Bergmann y Zech, 1994); no obstante, la información sobre su nutrición es relativamente poca y su disponibilidad limitada (Murillo y Alvarado, 2012, Stuhrmann et al., 1994). Algunas investigaciones informan que las concentraciones de nutrimentos en melina varían según la intensidad lumínica (Evans, 1979), la edad fisiológica de las hojas y su posición en la copa (Evans, 1979; Sampaio y Huber, 1999), y el orden de suelo (González, 1996). De acuerdo con Evans (1979), la posición de las hojas en la copa y la edad fisiológica del tejido se encuentran dentro de los principales factores que causan variaciones en las concentraciones foliares de G. arborea. Variables como la edad del árbol y la época de muestreo foliar no han sido analizadas en la melina (Evans, 1979; Sampaio y Huber, 1999; Stuhrmann et al., 1994). El efecto de la edad sobre las concentraciones de nutrimentos de esta especie se ha dirigido, principalmente, a la edad fisiológica de la hoja (Evans, 1979; Sampaio y Huber, 1999).
Varios autores (Boardman, Cromer, Lambert y Webb, 1997; Drechsel y Zech, 1991; Murillo y Alvarado, 2012; Stuhrmann et al., 1994) han determinado rangos adecuados de concentración foliar de nutrimentos y niveles críticos de concentración foliar en melina; sin embargo, la información aún no se considera suficiente para el caso de Costa Rica. Por lo anteriormente mencionado, resulta importante realizar un estudio sobre la variación de la concentración de nutrimentos a nivel foliar en esta especie y estandarizar un procedimiento de muestreo foliar, donde se observen, cuidadosamente, las fuentes de variabilidad que pueden permitir la determinación de los niveles de concentraciones de los nutrimentos foliares para mejorar las condiciones de su desarrollo.
Este estudio se realizó con el objetivo de determinar el efecto de la época de muestreo, la edad del árbol y la posición de las hojas en la copa, sobre la concentración foliar de nutrimentos de melina. Esta información se utilizó para generar un procedimiento de muestreo foliar y los rangos adecuados de concentración foliar de macro y micronutrimentos para plantaciones de G. arborea de uno a 12 años, ubicadas en los cantones de Golfito, Corredores y Osa, en el Pacífico Sur de Costa Rica.
2. Metodología
2.1. Sitio de estudio
El estudio se realizó en la Zona Sur de Costa Rica, en los cantones de Osa, Golfito y Corredores; en las zonas de vida de bosque muy húmedo premontano transición a basal y bosque muy húmedo tropical, de acuerdo con la clasificación de Holdridge (Bolaños y Watson, 1993). El clima es cálido, de húmedo a muy húmedo (Hernández, 2005), con una precipitación promedio anual cercana a 4.000 mm y una época seca que va de enero a marzo (IMN, 2008). Según Alvarado (2012), Palmar y el Valle de Coto tienen una precipitación promedio anual de 3.400 mm y 4.000 mm anuales, respectivamente; mientras que en Piedras Blancas caen 4.972 mm anuales. Esta zona tiene una temperatura máxima de 33,5 ºC, una mínima de 22 ºC y una media anual de 27 ºC y presenta las máximas temperaturas en el mes de marzo. Las plantaciones se encuentran ubicadas, en su mayoría, sobre suelos inceptisoles, que son suelos con un desarrollo moderado, en suelos entisoles con poco desarrollo de horizontes y con un exceso de humedad, y suelos ultisoles, considerados como suelos bien desarrollados, generalmente profundos, bien drenados y de color rojo y amarillo (Ortiz y Soto, 2008).
2.2. Muestreo foliar
Se seleccionaron doce plantaciones con topografía plana, con una pendiente no mayor al 5%, con edades de 1; 2; 4; 5; 5,5; 6; 6,5; 7; 7,5; 8,5; 9 y 12 años, ubicadas en los cantones de Osa, Golfito y Corredores. Se seleccionaron dos árboles dominantes por edad, se muestrearon en la época lluviosa y en la época seca de la zona (menos lluviosa).
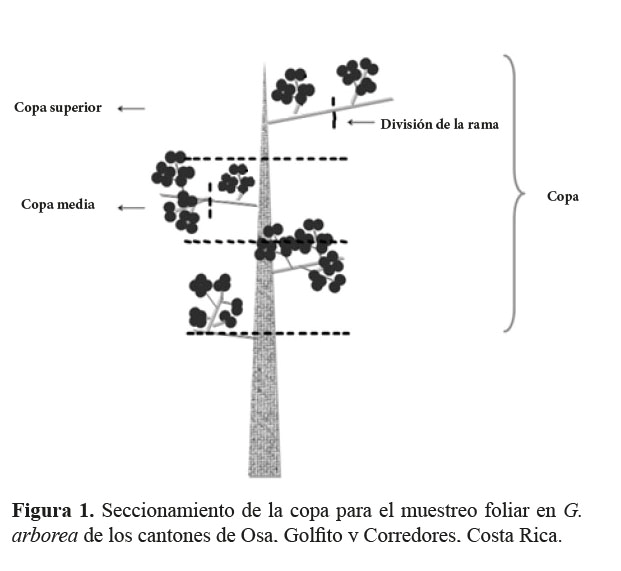
Estos árboles dominantes presentaron los mayores diámetros y alturas, buen estado fitosanitario (sin presencia de síntomas visibles de deficiencia nutricional) y una calidad de fuste mayor a 6 según Zeaser (1993). Se cortaron 45 árboles en total: 23 árboles en la época lluviosa y 22 árboles en la época seca. Las diferencias entre el número de árboles se deben a que para la edad de 6 años, en época lluviosa, solo se localizó un árbol, mientras que en la época seca, para la edad de 7,5, no se encontraron árboles con las características requeridas. De cada par de árboles dominantes por edad y para cada época, se tomó una muestra compuesta (dos árboles) de cada posición de la copa: una de la copa superior externa y otra de la copa superior interna y, por aparte, otra muestra de la copa media externa y otra de la copa media interna. Para seccionar la copa se midió la distancia desde el punto en el que se encontraron hojas hasta el ápice del fuste y esta se dividió en tres partes proporcionales: la copa superior, copa media y copa baja. Cada rama de la copa superior y de la copa media se seccionó en dos partes iguales: parte externa (extremo superior) y parte interna (extremo inferior) (figura 1) y de cada parte de la rama se muestreó el tercer par de hojas maduras con aspecto saludable. Cada muestra compuesta consistió de 30 hojas (15 hojas de cada uno de los dos árboles recolectados por edad y época). Se obtuvo un total de 91 muestras foliares, 48 muestras en la época lluviosa y 43 muestras en la época seca. De las 48 muestras de la época lluviosa, 12 muestras fueron de la copa superior externa, 12 de la copa superior interna, 12 de la copa media externa y 12 de la copa media interna. De las 43 muestras de la época seca, 11 fueron de la copa superior externa, 10 de la copa superior interna, 11 de la copa media externa y 11 de la copa media interna.
Cada muestra se almacenó en bolsas de papel con su respectiva identificación y se enviaron al Laboratorio de Suelos y Foliares del Centro de Investigaciones Agronómicas de la Universidad de Costa Rica, donde determinaron el N por combustión seca en un (Dumas) auto-analizador por incineración a 500 oC; el P, Ca, Mg, K, S, Fe, Cu, Zn, Mn, B y Al por digestión húmeda con HNO3 y determinación por espectrofotometría de emisión atómica con plasma (Kalra, 1998).
2.3. Análisis estadístico
Se realizó una prueba t-Student con una probabilidad de 5% de error para comparar las concentraciones foliares de macro y micronutrimentos obtenidas en la época lluviosa con las obtenidas en la época seca sin distinción de la posición de copa y edad del árbol. Se realizó un análisis de varianza para comparar las concentraciones foliares de macro y micronutrimentos de las cuatro posiciones de la copa tanto para la época lluviosa como para la época seca sin distinción de edad del árbol. Se realizó una prueba de Tukey en la que se encontraron diferencias significativas. Para evaluar la variación en las concentraciones foliares de los nutrimentos debidas a la edad del árbol, se agruparon las concentraciones de macro y micronutrimentos en tres categorías etáreas: 1-4, 5-8 y 9-12 años y se compararon por un análisis de varianza, con una probabilidad de 5% de error, sin distinción de posición de la copa tanto en la época lluviosa como en la época seca. En los casos en los que se encontraron diferencias significativas, se utilizó la comparación de medias múltiples de Tukey. Se usó el programa estadístico InfoStat 2.1. Debido a que la cantidad de muestras no estaban balanceadas, se hizo uso del método de “corrección de Satterthwaite” en todas las pruebas de “T de student” realizadas, el cual automáticamente corrige los cálculos, si las varianzas no son homogéneas. Para la determinación de los rangos adecuados de concentración, se tomaron los datos de las concentraciones foliares de macro y micronutrimentos de la época lluviosa y se calcularon los intervalos de confianza al 95%. Se seleccionaron las muestras de la época lluviosa porque, en esta, los árboles presentan la mayor demanda de nutrimentos y la mayor tasa de crecimiento. A partir de los resultados de la variación de las concentraciones foliares obtenidos en este estudio y a la consulta de fuentes bibliográficas, se determinó un protocolo preliminar de muestreo para G. arborea.
3. Resultados y discusión
3.1. Efecto de la época lluviosa y seca sobre las concentraciones foliares de nutrimentos en G. arborea
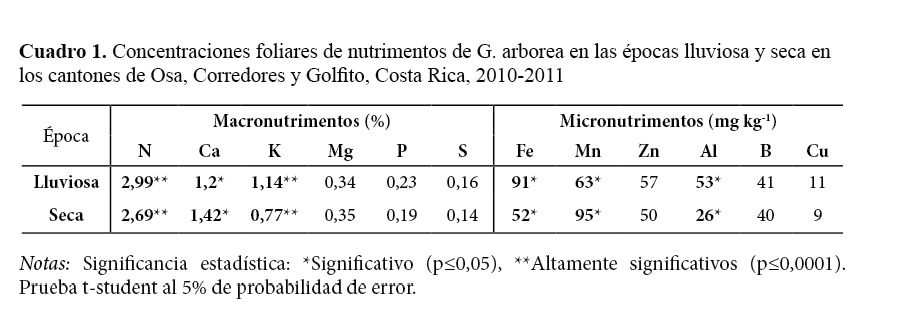
Al comparar las concentraciones foliares de los macronutrimentos en ambas épocas (Cuadro 1) se encontró que el mayor valor observado fue el N (2,99% y 2,69%), seguido por Ca (1,20% y 1,42%) y K (1,14% y 0,77%). Los otros macronutrimentos mostraron concentraciones inferiores a 0,34%, siendo el S el macronutrimento con menor porcentaje dentro del tejido foliar (0,16% y 0,14%). El orden de mayor a menor concentración foliar de los macronutrimentos en ambas épocas fue N>Ca>K>Mg>P>S. En los micronutrimentos el Fe (91 y 52 mg kg-1), Mn (63 y 95 mg kg-1) y el Zn (57 y 50 mg kg-1) presentaron las mayores concentraciones en ambas épocas, mientras que el Al (53 y 26 mg kg-1), B (41 y 40 mg kg-1) y Cu (11 y 9 mg kg-1) presentaron las menores concentraciones. El orden de mayor a menor concentración de los micronutrimentos varió entre épocas. En la época lluviosa correspondió a Fe>Mn>Zn>Al>B>Cu, mientras que en la época seca el orden fue Mn>Fe>Zn>B>Al>Cu (Cuadro 1).
Se observaron diferencias altamente significativas (p ≤ 0,0001) entre las concentraciones obtenidas en ambas épocas para los nutrimentos N y K y diferencias estadísticamente significativas (p ≤ 0,01) para el Ca, Fe, Mn y Al (Cuadro 1). Al respecto algunos autores (Leaf, 1973; Boardman et al., 1997; Smith y Loneragan, 1997) indican que los cambios estacionales provocan variaciones en las concentraciones foliares. Los nutrimentos N, K, Fe y Al presentaron mayores concentraciones foliares en la época lluviosa respecto de la seca (Cuadro 1). Comportamiento diferente presentaron el Ca y el Mn, cuyas concentraciones disminuyeron en un 18% y 51% en la época lluviosa respecto de la época seca. Mead (1984) indica que las concentraciones de los nutrimentos móviles como N y K tienden a disminuir sus concentraciones en la época seca, mientras que los nutrimentos menos móviles como el Ca se acumulan en los tejidos a lo largo de la época de crecimiento. Resultados similares obtuvieron Segura, Castillo, Alvarado y Blanco (2006) en A. acuminata, los que indican que las bajas concentraciones de Mn en la época lluviosa se deben a un efecto de dilución, pues en esta época el árbol entra en etapa de crecimiento y desarrollo de estructuras de almacenamiento; sin embargo, en la misma época, pero en suelos typic epiaquands, con bajos contenidos de Ca, Mg, K y P se puede dar una toxicidad a concentraciones foliares entre 380-640 mg kg-1 de Mn.
Rodríguez y Álvarez (2010) indican que las fluctuaciones en las concentraciones foliares debidas a la época son determinadas por factores ambientales como la temperatura y la humedad del suelo. Las menores concentraciones obtenidas en la mayoría de los nutrimentos de G. arborea en la época seca podrían estar asociadas a la poca humedad del suelo (Drechsel y Zech, 1993; van den Driessche, 1974; Nelson, 1989). Esto, pues según Pritchett (1986), la disponibilidad de agua reduce su crecimiento, lo que podría mermar la absorción de los nutrimentos. En regiones templadas, hay variaciones de concentración foliar, según la época de muestreo, que obligan a definirla, tratando de minimizar los costos de este tipo de análisis (Vidal, Ferrada y Riquelme, 1984).
3.2. Efecto de la posición de las hojas en la copa de los árboles sobre la concentración foliar de nutrimentos en G. arborea según la época de muestreo
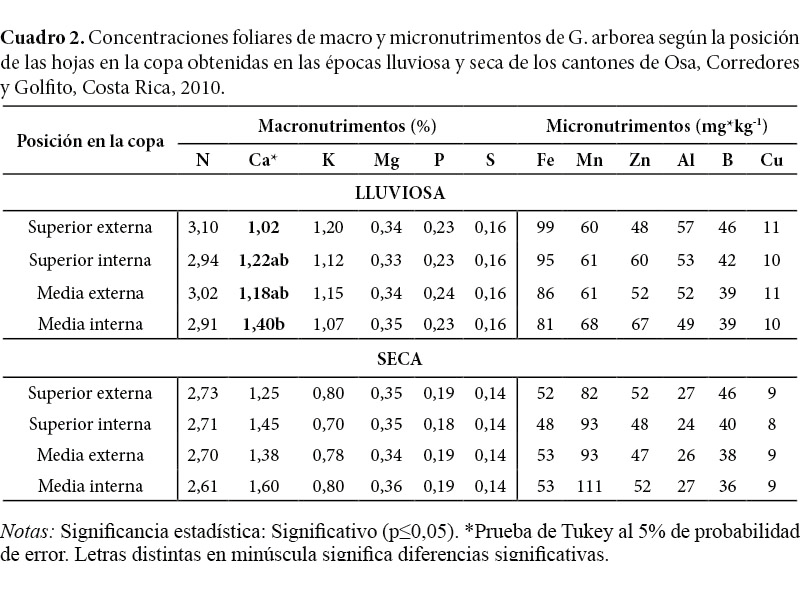
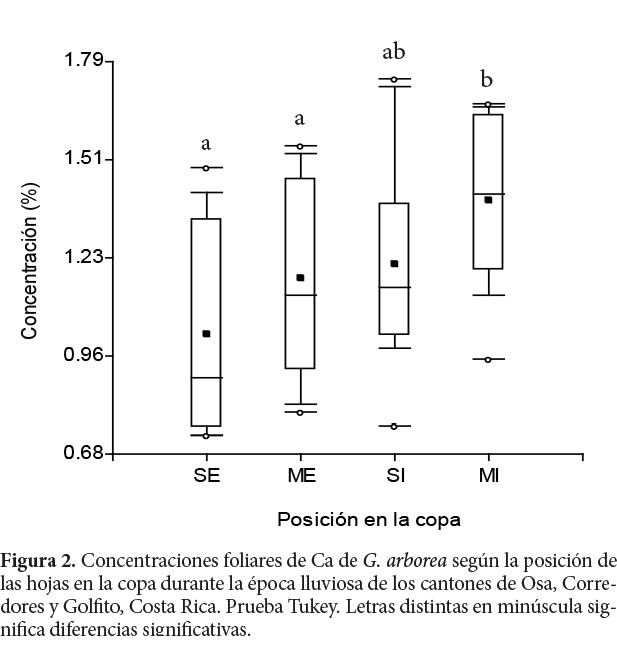
De acuerdo con Leaf (1973) y Evans (1979), las concentraciones foliares de nutrimentos varían a nivel vertical y horizontal en la copa. Para Drechsel y Zech (1993), dos tendencias influyen en este comportamiento: el aumento de la sombra de arriba hasta abajo y de exterior a interior de la copa. En el caso de la melina, este comportamiento no se presentó, pues en la mayoría de los nutrimentos no se encontraron diferencias significativas debidas a la posición de las hojas en la copa en ambas épocas de muestreo. Únicamente la concentración de Ca en la época lluviosa presentó mayores concentraciones (p=0,02) en la copa media interna, en un 27% mayor que las obtenidas en la copa superior externa (Cuadro 2 y Figura 2). Según Onyekwelu, Mosandl y Stimm, (2006), el Ca se almacena en las hojas viejas, aumentando la concentración en estas hasta su senescencia cuando, al caer, incorporan este nutrimento al suelo como se ha documentado en plantaciones de teca (Márquez, Hernández, Torres y Franco, 1993; Nath, Banerjee, Chattotaj, Ganguly, Das y Banerjee, 1988; Samndi y Jibrin, 2012) y de A. mangium (Garay, Kindel, Carneiro, Franco, Barroa y Abbade, 2003). Este comportamiento de Ca merece atención, principalmente, en suelos con baja disponibilidad de este mismo (Sampaio y Huber, 1999).
Aunque no significativo, ciertos macronutrimentos y micronutrimentos, en ambas épocas, presentaron mayores concentraciones en alguna de las partes de la copa. En la época lluviosa, las concentraciones de N, K, Fe, Al y B tendieron a ser mayores en la copa superior externa. Mientras que las concentraciones de Mn y Zn tendieron a aumentar en la copa media interna (Cuadro 2). En la época seca no se encontraron patrones de concentración de los nutrimentos tan definidos en las partes de la copa como en la época lluviosa, a excepción del Ca y Mn que se concentraron en mayor cantidad en la copa media interna y el B que tendió a ser mayor en la copa superior externa (Cuadro 2). Según Smith y Loneragan (1997), las concentraciones de nutrimentos móviles como el N aumentan en los tejidos nuevos del árbol (copa superior externa), debido a que las hojas nuevas se desarrollan a expensas de la reserva de estos nutrimentos presentes en las hojas viejas. Por el contrario, los nutrimentos inmóviles como el Ca aumentan en los tejidos viejos (copa media interna) donde quedan capturados debido a su poca movilidad (Sampaio y Huber, 1999).
Segura et al. (2006) atribuyen el poco efecto de la copa sobre las concentraciones foliares, en el jaúl, a factores como: pocas ramas en la copa (en particular en plantaciones viejas muy densas), el tamaño de las hojas, la pérdida de follaje, la autopoda de la especie, la poda y raleo de los árboles, e indica que estos factores permiten un nivel de luminosidad similar en las hojas durante todo el año, lo que mantiene el árbol fotosintéticamente activo y disminuye el efecto de la posición de la copa. Diferentes resultados fueron obtenidos por Sampaio y Huber (1999) en plantaciones de melina de la región de Manaos, donde encontraron mayores concentraciones de K en la parte superior de la copa y mayores concentraciones de Ca en la parte media de la copa. Evans (1979) halló que las concentraciones de N, P, Zn y B disminuyen y las concentraciones de Ca y Fe aumentan con el incremento de la sombra en la copa. En otras especies como el jaúl (Segura et al., 2006) se han informado variaciones en las concentraciones de nutrimentos debidas a la posición de las hojas en la rama.
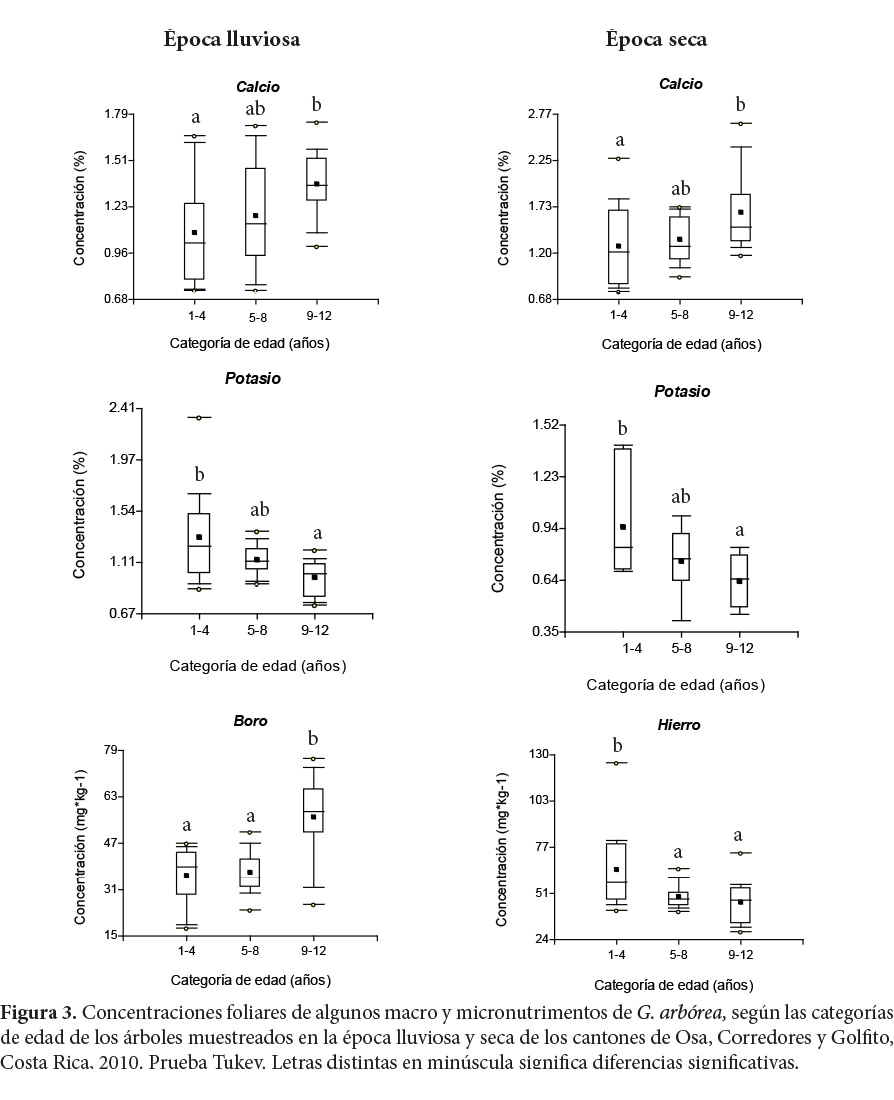
3.3. Variación de las concentraciones foliares de nutrimentos en G. arborea debido a la edad de los árboles por época de muestreo
En la época lluviosa la concentración foliar de B de edades entre 1-4 y 5-8 fue similar, pero significativamente menor que la concentración del grupo etario 9-12 (p ≤ 0,0001), la cual en estos árboles fue mayor en un 36% respecto a los árboles de uno a cuatro años. Tanto en la época húmeda como en la seca, la concentración foliar de Ca entre los grupos etarios 1-4 y 5-8 fue menor a la del grupo entre 9-12 años (p ≤ 0,05); el Ca presentó concentraciones mayores en un 21% en árboles de 9-12 años respecto a árboles de uno a cuatro años. La concentración foliar de K fue mayor en el grupo etario 1-4 (p ≤ 0,05) que los grupos 5-6 y 9-12 en ambas épocas de muestreo, valores mayores en un 25% en relación con árboles de 9-12 años (Figura 3).
En la época seca se obtuvieron variaciones estadísticamente significativas (p ≤ 0,05) en las concentraciones foliares de Ca, K y Fe debidas a la edad. El K y el Fe en árboles de 1-4 años presentaron mayores concentraciones en un 33% y 30% respecto a árboles de 9-12 años. Mientras que el Ca en árboles de nueve a 12 años obtuvo concentraciones superiores en un 23% respecto a árboles de uno a cuatro años (Figura 3). El B en árboles de nueve a 12 años presentó 32% de mayor concentración respecto a árboles de uno a cuatro años, aunque no significativa (p=0,0593).
Estas variaciones, debidas a la edad de los árboles, presentan dos tendencias: los nutrimentos móviles como el K disminuyeron con la edad, mientras que los inmóviles como el Ca y la excepción del Fe y los parcialmente móviles como el B (Lehto, Ruuhola y Dell, 2010) tendieron a aumentar con la edad. Según Bertsh (1998), el Ca es un nutrimento que proporciona rigidez a las paredes celulares y conforme aumenta el crecimiento requiere mayor solidez, por lo que aumenta la demanda de este nutrimento. Un comportamiento muy similar al obtenido en el K lo encontraron Siddiqui, Shah y Yaqoob (2009) en teca, en Malasia, donde la concentración foliar de K presentó las mayores concentraciones en edades tempranas y tendió a disminuir con el aumento de la edad.
De acuerdo con Onyekwelu et al. (2006), en plantaciones de melina, en Nigeria, la edad del árbol es una variable que no afecta significativamente la concentración de nutrimentos foliares. Mills y Jones (1996, citados por Segura et al., 2006) indican que la poca variación de las concentraciones debidas a la edad puede ser efecto, con los años, de una adecuada disponibilidad de los nutrimentos en el suelo.
3.4. Rangos de concentración foliar de nutrimentos durante la época lluviosa para G. arborea de los cantones de Osa, Golfito y Corredores de Costa Rica
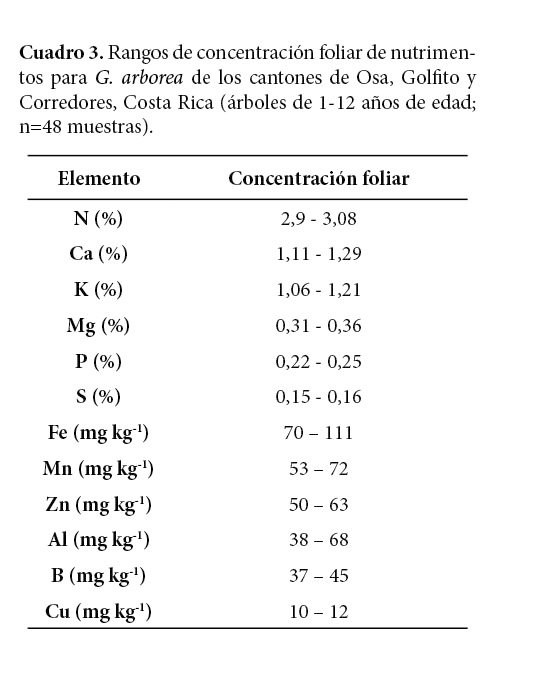
Los rangos de concentración foliar de nutrimentos para G. arborea se determinaron para cada macro y micronutrimento de la época lluviosa (Cuadro 3). Los rangos de concentraciones foliares de N, P, K, Ca, Mg, Fe, Mn, B, Zn y Cu del presente estudio se encuentran dentro de los valores considerados como adecuados por Murillo y Alvarado (2012). Excepciones a lo mencionado las constituyen el S, cuyas concentraciones se consideran como adecuadas en el presente estudio (0,15-0,16%) y como altas por Murillo y Alvarado (2012), y las del Al (38-68 mg kg-1) que están por debajo de los establecidos como adecuados por los autores anteriormente citados. Los niveles de concentración de Stuhrmann et al. (1994) para melina se hallan por debajo de los determinados en esta investigación –a excepción del Mn–, posiblemente debido a que las plantaciones se encuentran ubicadas en pendientes, en suelos ácidos y erosionados, con baja disponibilidad de N y P (Stuhrmann et al., 1994), características de sitios con factores limitantes para el desarrollo de la especie (Rojas y Murillo, 2004).
Algunas investigaciones han establecido recomendaciones para estandarizar el muestreo foliar en especies forestales (Brunck, 1987). Se consideran como recomendaciones generales las de Evans (1979), quien recomienda tomar el tercer par de hojas maduras en la parte superior de la copa de árboles sin flores ni frutos y con buena copa, en plantaciones de melina en Nueva Guinea o las de Drechsel y Zech (1991); Boardman et al. (1997); Reuter, Robinson, Peverill, Price y Lambert (1997), y Rodríguez y Álvarez (2010), quienes mencionan que en especies forestales se muestrean, principalmente, las hojas maduras de la copa superior del árbol. Para efectos de este estudio se sugiere el siguiente método preliminar de muestreo foliar para G. arborea.
3.5.1. Selección de los árboles
Se deben seleccionar árboles que tengan un crecimiento vigoroso (con diámetros y alturas similares), que sean dominantes, debido a que son representativos del sitio, con menores variaciones de nutrimentos entre árboles (Leaf, 1973) y poco sensibles a la sombra (Drechsel y Zech, 1993).
No se deben muestrear árboles:
- En época de floración o fructificación (Drechsel y Zech, 1993; Reuter et. al., 1997).
- Enfermos, con daños mecánicos o ataque de insectos u hongos (Drechsel y Zech, 1993; Jones y Steyn, 1973; Nelson, 1989, Reuter et al., 1997).
- Contaminados con polvo (Drechsel y Zech, 1993).
- Afectados por bajas temperaturas, exceso o deficiencia de humedad y estrés de temperatura (Jones y Steyn, 1973; Nelson, 1989; Reuter et al., 1997).
- En áreas con características inusuales como zonas rocosas, mal drenaje, cercanas a canales de agua y caminos, entre otras (Reuter et al., 1997).
3.5.2. Número de árboles, tipo de muestra y recolección
Se debe colectar material foliar de dos a cuatro árboles dominantes de melina (dependiendo del tamaño de los árboles); representativos del sitio, de forma tal que se asegure una muestra de hojas maduras que pesen un kilogramo de material fresco, aproximadamente 30 hojas. El material colectado del total de árboles muestreados se debe mezclar para formar una muestra compuesta y así disminuir la variabilidad entre árboles (Reuter et al., 1997). Para relacionar los valores de concentración foliar de nutrimentos, resulta ideal contar con valores de crecimiento de los árboles, por lo que se recomienda tomar las muestras más cercanas, en lo posible, a las parcelas permanentes de muestreo y, de ser viable, muestrear el suelo.
Se recomienda muestrear el tercer par de hojas maduras, bien desarrolladas, en árboles sin flores ni frutos (Evans, 1979). El tejido debe ser de color verde, de árboles sanos y con una buena iluminación (Boardman et al., 1997). No muestrear material vegetal muerto, dañado por insectos o lesiones mecánicas (Nelson, 1989).
El tejido se puede muestrear tanto en la copa media interna o externa como en la copa superior externa o interna del árbol, pues no se encontraron diferencias significativas en las concentraciones foliares debidas a las posiciones de la copa. No obstante, en aquellos en los que el acceso a la copa superior se vea limitado, se recomienda colectar material en la copa media externa.
Las hojas colectadas se deben depositar en bolsas de papel (Drechsel y Zech, 1993; Nelson, 1989), para evitar el desarrollo de hongos. Se deben identificar el número de árbol, la posición de la copa en la que se colectó la muestra, el nombre de la finca, la especie y el día de muestreo. De ser posible, es conveniente geo-referenciar el sitio de muestreo.
3.5.3. Época de muestreo
En regiones de clima monzónico (período seco/lluvioso) se recomienda colectar las muestras en el momento en que el árbol haya desarrollado su máximo crecimiento anual, normalmente coincidente con el establecimiento pleno de la época lluviosa (un mes después de la entrada de la lluvia); cuando el patrón de distribución de la lluvia es unimodal, se pueden tomar muestras en cualquier tiempo, siempre que no coincida con la floración, la fructificación o un período de defoliación fuerte. En la época lluviosa se encontrarán las mayores concentraciones de N, K, Fe y Al, y las concentraciones de P, S, Mg, Zn, Cu y B no disminuirán significativamente.
3.5.4. Almacenamiento y transporte
El transporte al laboratorio debe hacerse lo más rápidamente posible (Jones y Steyn, 1973); por tanto, se recomienda colectar la muestra temprano en la mañana (Reuter et al., 1997). En caso de que las muestras se tengan que mantener varios días en el sitio de trabajo antes de ser entregadas al laboratorio, se deben almacenar bajo condiciones de refrigeración o de aire acondicionado. Si no se contara con refrigeración, las muestras de melina manteniendo abiertas las bolsas de papel en sitios con buena ventilación pueden pasar un máximo de cuatro días bajo temperatura ambiente. El cuarto día es el límite para entregarlas al laboratorio. Al pasar un día más, las hojas empiezan a cambiar su color.
Las muestras no se deben transportar en compartimentos cerrados a temperatura ambiente, debido a que la temperatura se concentra y activa la respiración celular, alterando las concentraciones foliares de los nutrimentos. Lo más recomendado es transportarlas en condiciones de refrigeración; de lo contrario, se deben trasladar en bolsas de papel abiertas, de forma tal que la muestra se mantenga fresca.
4. Conclusiones
Los rangos de concentración foliar de nutrimentos de este estudio son una guía para el diagnóstico nutricional de plantaciones de melina establecidas en los cantones de Osa, Golfito y Corredores de Costa Rica. La época de muestreo (período seco o lluvioso) causó variaciones significativas en las concentraciones foliares de N, Ca, K, Al, Fe y Mn. Se obtuvieron mayores concentraciones foliares de N, K, Fe y Al en la época lluviosa; mientras que en la seca se lograron concentraciones mayores de Ca y Mn. Únicamente las concentraciones foliares de Ca en la época lluviosa variaron significativamente, según la posición de las hojas.
La edad de los árboles de melina causó variaciones significativas en las concentraciones de Ca, K, B y Fe. Los árboles maduros (edades de 9-12 años) presentan mayores concentraciones foliares de Ca y B que los árboles jóvenes (1-4 años). Los árboles jóvenes (1-4 años) mostraron una mayor concentración de K foliar en comparación con los árboles de edades entre 9 y 12 años. Se recomienda hacer los muestreos para el análisis foliar de melina cuando se hayan estabilizado las lluvias y colectar el tercer par de hojas de la copa media externa con buena iluminación.
5. Referencias
Aldrich, S. (1973). Plant Analysis: Problems and Opportunities. En: L. M. Walsh y J. D. Beaton (Eds), Soil Testing and Plant Analysis (pp. 223-248). Wisconsin, US: Soil Science Society of America.
Arce, H. y Barrantes, A. (2006). La madera en Costa Rica: Situación actual y perspectivas. Recuperado de http://www.oficinaforestalcr.org/media_files/download/MADERAEN COSTARICA,SITUACIONACTUALYPERSPECTIVAS.2006.pdf
Alvarado, A. (2012). Suelos de Costa Rica: Origen y caracterización de los suelos del Pacífico Central y Sur (Primer borrador). San José, Costa Rica: Centro de Investigaciones Agronómicas, Universidad de Costa Rica.
Bertsh, F. (1998). La fertilidad de los suelos y su manejo. San José, Costa Rica: ACCCS.
Binkley, D. (1993). Nutrición forestal: Prácticas de manejo. Distrito Federal, México: Editorial Mexicana.
Boardman, R., Cromer, R., Lambert, M. y Webb, M. (1997). Forest Plantations. En D. J. Reuter y J. B. Robinson (Eds), Plant Analysis and Interpretation Manual (pp. 503-572). AU: CSIRO Publishing.
Bolaños, R. y Watson, V. (1993). Mapa ecológico de Costa Rica. (Según clasificación de Zonas de vida del mundo de L. R Holdridge). Escala. Colores. 1:200000. San José, Costa Rica: Instituto Geográfico Nacional.
Brockley, R. (2001). Foliar analysis as a planning tool for operational fertilization. Proceedings of Enhanced Forest Management: Fertilization & Economics Conference. Recuperado de http://www.for.gov.bc.ca/hre/standman/docs/10Brockley.PDF
Brunck, F. (1987). Tropical Forest Species. En P. Martin-Prével, J. Gagnard y P. Gautier (Eds), Plant analysis as a guide to the nutrient requeriments of temperate and tropical crops (pp. 479-486). New York: Lavoisier Publishing.
Chijioke, E. (1980). Impact on soils of fast growing species in lowland humid tropics. FAO Forestry Paper 21. Roma: FAO.
Correndo, A. y García, F. (2012). Concentración de nutrimentos en planta como herramienta de diagnóstico: Cultivos extensivos. Recuperado de http://lacs.ipni.net/article/LACS-1155.
Drechsel, P. y Zech, W. (1991). Foliar nutrient levels of broad-leaved tropical trees: A tabular review. Plant and soil, 131(1), 29-46.
Drechsel, P. y Zech, W. (1993). Mineral Nutrition of Tropical Tress. En L. Pancel (Ed), Tropical Forestry Handbook (p. 515-567). Hamburg, Springer-Verlag.
Evans, J. (1979). The effects of leaf position and leaf age in foliar analysis of Gmelina arborea. Plant and soil, 52(4), 547-552. Recuperado de http://dx.doi.org/10.1007/BF02277950
Garay, I., Kindel, A., Carneiro, R., Franco, A., Barroa, E. y Abbade, L. (2003). Comparação da materia organica e de outros atributos do solo ente plantações de Acacia mangium e Eucalyptus grandis. Revista Brasileira. Ciência do Solo, 27, 705-712. Recuperado de http://dx.doi.org/10.1590/S0100-06832003000400015
González, E. (1996). Tropical tree species for reforestation: Studies on seed storage foliar nutrient content and wood variation (Tesis de doctorado). Texas A&M University, Texas.
Hernández, G. (2005). Mapa climatológico de Costa Rica (CD-ROOM), Escuela de Ciencias Geográficas de la Universidad Nacional, Heredia, Costa Rica.
IMN (2008). Clima, variabilidad, cambio climático en Costa Rica. Recuperado de http://www.slideshare.net/Socialesdigital/el-clima-variabilidad-y-cambio-climatico-en-cr.
Jones, J. y Steyn, W. (1973). Sampling, Handling, and Analyzing Plant Tissue Samples. En L. M. Walsh y J. D. Beaton (Eds), Soil Testing and Plant Analysis (pp. 249-270). Wisconsin: Soil Science Society of America.
Kalra, Y. (1998). Handbook of reference methods for plant analysis. Boca Raton, Florida: Soil and Plant Analysis Council.
Leaf, A. (1973). Plant analysis as an Aid in Fertilizing Forest. En L. M. Walsh y J. D. Beaton (Eds), Soil Testing and Plant Analysis (pp. 427-454). Wisconsin: Soil Science Society of America.
Lehto, T., Ruuhola, T. y Dell, B. (2010). Boron in forest trees and forest ecosystems. Forest Ecology and Management, 260, 2053-2069. Recuperado de http://dx.doi.org/10.1016/j.foreco.2010.09.028
Márquez, O., Hernández, R., Torres, A. y Franco, W. (1993). Cambios en las propiedades físico-químicas de los suelos en una cronosecuencia de Tectona grandis. Turrialba, 43(1), 37-41.
Mead, D. (1984). Diagnosis of nutrient deficiencies in plantations. En E. Nambiar y G. Bowen (Eds), Nutrition of plantation forests (pp. 259-291). London: Academic Press.
Munson, R. y Nelson, W. (1973). Principles and Practices in Plant Analysis. En L. M. Walsh y J. D. Beaton (Eds), Soil Testing and Plant Analysis (p. 223-248). Wisconsin: Soil Science Society of America.
Murillo, R. y Alvarado, A. (2012). Nutrición y fertilización de Gmelina arbórea (Capítulo 15). En A. Alvarado y J. Raigosa (Eds), Nutrición y fertilización forestal en regiones tropicales (pp. 285-298). San José, Costa Rica: Centro de Investigaciones Agronómicas, Universidad de Costa Rica.
Nath, S., Banerjee, M., Chattotaj, G., Ganguly, S., Das, P. y Banerjee, S. (1988). Changes in soil attributes consequent upon differences in forests cover in a plantation area. Journal of the Indian Society of Soil Science, 36, 515-521.
Nelson, W. (1989). Plant and tissue testing principles relating to the identification of nutrient deficiencies. En D. L. Plucknett y H. B. Sprague (Eds), Detecting Mineral Nutrient Deficiencies in Tropical and Temperate Crops (pp. 23-40). US: Westview Press, International Book Distributors.
Onyekwelu, J., Mosandl, R. y Stimm, B. (2006). Productivity, site evaluation and state of nutrition of Gmelina arborea plantations in Oluwa and Omo forest reserves, Nigeria. Forest Ecology and Management, N.° 229, 214–227. Recuperado de www.elsevier.com/locate/foreco. http://dx.doi.org/10.1016/j.foreco.2006.04.002
Ortiz, E. y Soto, C. (2008). Atlas digital de Costa Rica (1CD-ROOM). Cartago, Costa Rica: Laboratorio de Sistemas de Información Geográfica, Escuela de Ingeniería Forestal. Instituto Tecnológico de Costa Rica.
Pritchett, W. (1986). Suelos forestales, propiedades, conservación y mejoramiento. México: México-LIMUSA.
Reuter, D., Robinson, J., Peverill, K., Price, G. y Lambert, M. (1997). Guidelines for Collecting, Handling and Analysing Plant Materials. En D. J. Reuter y J. B. Robinson (Eds), Plant Analysis and Interpretation Manual. AU: CSIRO Publishing.
Rojas, F. y Murillo, O. (2004). Botánica y ecología. En F. Rojas et al. (Eds), Manual para productores de melina (Gmelina arborea) en Costa Rica. Cartago, Costa Rica.
Rodríguez, J. y Álvarez, J. (2010). Nutrición y fertilización de plantaciones forestales. Santiago, Chile: Gráfica LOM.
Samndi, A. y Jibrin, J. (2012). Pedogenesis and classification of soils under teak (Tectona grandis Linn. f) plantation of various ages in the southern Guinea savanna of Nigeria. Asian Journal of Agricultural Sciences, 4(1), 16-25.
Sampaio, L. y Huber, W. (1999). Concentração e distribuição de nutrimentos nas folhas de espécies florestais, na Amazônia Ocidental. Floresta e Ambiente, 6(1), 127-137.
Segura, M., Castillo, A., Alvarado, A. y Blanco, F. (2006). Variación del contenido foliar de nutrimentos de Alnus acuminata. Agronomía Costarricense, 30(1), 53-63.
Siddiqui, M., Shah, A. y Yaqoob, S. (2009). Chronosequence and crown strata effects on foliar nutrient concentrations in teak (Tectona grandis L.f) Plantations. Pak. J. Bot., 41(6), 3023-3034.
Smith, F. y Loneragan, J. (1997). Interpretation of Plant Analysis: Concepts and Principles. En D. J. Reuter y J. B. Robinson (Eds), Plant Analysis and Interpretation Manual. AU: CSIRO Publishing.
Stuhrmann, M., Bergmann, C. y Zech, W. (1994). Mineral nutrition, soil factors and growth rates of Gmelina arborea plantations in the humid lowlands of northern Costa Rica. Forest Ecology and Management, N.° 70, 135-145. Recuperado de http://dx.doi.org/10.1016/0378-1127(94)90081-7
van den Driessche, R. (1974). Prediction of mineral nutrient status of trees by foliar analysis. The Botanical Review, 40(3),347-396. Recuperado de http://dx.doi.org/10.1007/BF02860066
Vidal, I., Ferrada, R. y Riquelme, E. (1984). Evolución estacional de nutrimentos en Pinus radiata D. Don en Chile. Turrialba, 34(3), 261-266.
Zeaser, D. (1993). Evaluación de árboles de melina: Clasificación de forma, sistema DANIDA modificado. Departamento de Investigación y desarrollo, STON Forestal S.A. S.l.

Revista de Ciencias Ambientales por Universidad Nacional de Costa Rica se distribuye bajo una Licencia Creative Commons Atribución 4.0 Internacional.
Basada en una obra en http://www.revistas.una.ac.cr/ambientales.
Permisos que sobrepasen el ámbito de esta licencia, pueden ser consultados en revista.ambientales@una.cr