
Revista de Ciencias Ambientales (Trop J Environ Sci). (Diciembre, 2016). EISSN: 2215-3896. Vol 50(2): 1-19.
DOI: http://dx.doi.org/10.15359/rca.50-2.1
URL: www.revistas.una.ac.cr/ambientales
EMAIL: revista.ambientales@una.cr

Evaluación de la concentración de macro y micronutrimentos en la biomasa aérea (BA) de la melina (Gmelina arborea) de uno a 12 años de edad en la zona sur de Costa Rica
Evaluation of Macro and Micronutrients Concentration in the Melina (Gmelina arborea) Aerial Biomass (AB) of One to 12 Years Old in Southern Costa Rica
Rafael Murillo-Cruza, María José Avellán-Zumbadob, Carlos E. Ávila-Ariasc y
Alfredo Alvarado-Hernándezd
a Máster en Ingeniería Forestal con especialidad en suelos, investigador en el Instituto de Investigación y Servicios Forestales de la Universidad Nacional de Costa Rica, murillorafael5454@yahoo.com
b Licenciada en Ingeniería Forestal con especialidad en producción, investigadora en la Escuela de Ciencias Ambientales de Universidad Nacional de Costa Rica, maria.avellan.zumbado@una.cr
c Máster en Ingeniería Forestal con especialidad en producción, investigador en el Instituto de Investigación y Servicios Forestales de la Universidad Nacional de Costa Rica, carlosenriquea٧٩@gmail.com
d Doctor en Ingeniería Agrónoma con especialidad en suelos, profesor emérito en el Centro de Investigaciones Agronómicas de la Universidad de Costa Rica, alfredo.alvarado@ucr.ac.cr
[Recibido: 20 de abril 2016; Aceptado: 13 de junio 2016; Corregido: 26 de octubre 2016; Publicado: 30 de noviembre 2016]
Resumen
Se determinó la concentración de macro y micronutrimentos en los componentes (tronco, corteza, hojas, ramas primarias y secundarias) de la biomasa aérea (BA) de la melina de uno a 12 años de edad en la zona sur de Costa Rica y los intervalos de confianza de las concentraciones para N, P, K, Ca, Mg, S, Fe, Mn, Zn, Cu y B. El muestreo consistió de dos árboles por edad, excepto en una edad, por lo que se cortó un total de 23 árboles distribuidos en las edades de uno a 12 años. Se correlacionaron algunas variables del suelo (pH, acidez del suelo, SA y CICE) con la concentración de macro y micronutrimentos en los distintos componentes aéreos de la melina. El orden de los componentes de biomasa, según la concentración total de macronutrimentos, fue: las hojas (6,3%), la corteza (3,57%), las ramas secundarias (2,56%), las ramas primarias (1,72%) y el fuste con la menor concentración total (0,78%). Los macroelementos de mayor concentración individual en todos los componentes fueron el N, Ca y K y los menos abundantes, el Mg, P y S. El orden de los componentes según la concentración total de micronutrimentos fue la corteza (765 mg kg-1), las ramas primarias (323 mg kg-1), las ramas secundarias (314 mg kg-1), las hojas (295 mg kg-1) y el fuste (190 mg kg-1). La concentración de Ca aumenta en la corteza, ramas primarias y secundarias, mientras que la de K disminuye (en todos los componentes) conforme aumenta la edad de la plantación. En todos los componentes de la BA existe una correlación positiva de media a alta de la concentración de Fe y Mn con la acidez extraíble del suelo.
Palabras clave: Lamiaceae, nutrimentos, plantación forestal, suelo.
Abstract
Concentration of macro and micronutrients in one to 12 years old melina aboveground biomass (AB) components (stem, bark, leaves, primary and secondary branches) in the south zone of Costa Rica and confidence intervals of concentrations for N, P, K, Ca, Mg, S, Fe, Mn, Zn, Cu, and B were determined. Sampling consisted of two trees by age, with the exception of one age. Then, a total of 23 trees were cut and distributed in ages from one to 12 years. Some soil variables (pH, soil acidity, SA, and CICE) were correlated with the concentration of macro and micronutrients in the melina aerial components. The order of biomass components in accordance with total concentration of macronutrients was leaves (6.3%), bark (3.57%), secondary branches (2.56%), primary branches (1.72%), and the stem with the lowest total concentration (0.78%). The largest single concentration of macroelements in all components was the N, Ca, and K, and the least abundant was the Mg, P, and S. The order of the components in the total concentration of micronutrients was bark (765 mg kg-1), primary branches (323 mg kg-1), secondary branches (314 mg kg-1), leaves (295 mg kg-1), and the stem (190 mg kg-1). The concentration of Ca increases in the bark, primary, and secondary branches, while K decreases (in all components) with increasing age of the plantation. In all AB components there is a high positive correlation with average concentration of Fe and Mn with removable soil acidity.
Keywords: Forest plantation, Lamiaceae, nutrients, soil.
La melina (Gmelina arborea Roxb) pertenece a la familia Lamiaceae (Kok, 2012), es una de las especies más utilizadas en proyectos de reforestación en Costa Rica (Arce, 2012) y es muy exigente de suelos fértiles para su desarrollo productivo (Murillo, 1996, Agus et al., 2004). La determinación de la concentración de los nutrimentos en los componentes de la BA (biomasa aérea) de los árboles (tronco, corteza, ramas y hojas) de la melina es importante para la determinación de desórdenes nutricionales y deficiencias asintomáticas (Correndo y García, 2012), también para indicar interacciones entre nutrimentos e identificar zonas de las plantaciones con comportamiento diferente, en la planificación de la fertilización de las nuevas plantaciones (Ogbonnaya y Kinako, 1993), en el monitoreo del manejo nutricional de la plantación, el cálculo de los nutrimentos removidos en la cosecha (Agus et al., 2000b) y el aporte al reciclaje de nutrimentos, (Olaleye, Aderinola, Onasanya y Akinbola, 2002).
Algunos estudios han abordado el tema de la concentración en algunos componentes de la BA de la especie (Agus, Karyanto, Hardiwinoto, Haibara y Toda, 2000a; Arias, Calvo, Richter y Dohrenbusch, 2011; Negi, Bahuguna y Sharma, 1990). En general, se le ha dado mayor importancia a la investigación de los macronutrimentos (Negi et al., 1990) y menos a los micronutrimentos (Avellán, 2012; Evans, 1979). La variación de la concentración de nutrimentos en la BA de la melina ha sido estudiada según la posición de las hojas en la copa o en las ramas (Avellán, 2012; Evans, 1979; Sampaio y Huber, 1999), el estado fisiológico de los árboles (Evans, 1979; Osundina y Osonubi, 1989), el efecto de la edad (Avellán, 2012; Caguasango, 2012), el efecto del período de lluvias (Avellán, 2012), la posición sociológica en el dosel (Sampaio y Huber, 1999), la posición del árbol en la pendiente (Stuhrmann, Bergmann y Zech, 1994), la relación con los nutrimentos disponibles del suelo (Caguasango, 2012; Escobar, 2013) y comparación con otras especies (Arias et al., 2011; Sampaio y Huber, 1999).
La concentración de nutrimentos en los componentes aéreos de la melina varía según los diferentes componentes de la BA (Negi et al., 1990), pero no es significativamente diferente cuando cambia el diámetro del árbol (Agus, et al., 2004; Onyekwelu, Mosandl y Stimm, 2006) o la edad de las plantaciones entre 10 y 28 años (Onyekwelu et al., 2006). Según Negi et al., (1990), la concentración del Mg es mayor que la del P en los distintos componentes de la BA de la melina. El Ca es el elemento de mayor concentración en la corteza y las ramas primarias y secundarias; sin embargo, el K es el elemento de mayor concentración en el tronco (Negi et al., 1990).
El componente de la BA de la melina más estudiado ha sido las hojas. Varias investigaciones han estudiado la variación de la concentración foliar en melina (Agus et al., 2000b; Avellán, 2012; Caguasango, 2012; Dreschsel y Zech, 1991; Escobar, 2013; Evans, 1979; Sampaio y Huber, 1999; Stuhrmann et al., 1994). La concentración foliar de N, P, K, Mg, Zn tiene la tendencia a disminuir en las hojas más viejas, debido a la movilidad de estos elementos y; por el contrario, el Fe, Ca, Cu y B tienden a disminuir en hojas jóvenes por su poca movilidad dentro de la planta (Evans, 1979). En general, el orden de mayor a menor concentración foliar de macroelementos es N>K>Ca>Mg>S>P; sin embargo, Negi et al. (1990) reportaron el mismo orden, excepto por una mayor concentración foliar de Ca que de K. El orden de la concentración foliar de los microelementos es Fe>Mn>Zn (Sampaio y Huber, 1999). Según Correndo y García (2012), la concentración de macro y micronutrimentos en las plantas está estrechamente relacionada con la capacidad del suelo para suplirlos y otros autores (Osundina y Osonubi, 1989) consideran como importante la interacción entre el árbol, el ambiente y el manejo de la plantación.
El objetivo del artículo es determinar la concentración de macro y micronutrimentos en los componentes aéreos de la BA (tronco, corteza, hojas, ramas primarias y secundarias) de la melina, la tendencia de esta concentración de uno a 12 años en la zona sur de Costa Rica y su relación con algunas propiedades del suelo. Los resultados podrían utilizarse para mejorar el conocimiento de la nutrición de esta especie, y dirigir la investigación y la fertilización como parte de exitosos proyectos de reforestación.
2. Materiales y métodos
El estudio se llevó a cabo con árboles de melina recolectados en los cantones de Osa, Corredores y Golfito en la provincia de Puntarenas, Costa Rica, entre las coordenadas 8,06140_9,32680 latitud norte y -82,82980_-83,90210 longitud oeste, en las zonas de vida bosque muy húmedo Premontano transición a Basal y bosque muy húmedo Tropical, de acuerdo con la clasificación de Holdridge (Holdridge, 1982). La precipitación promedio anual es cercana a 4 000 mm con una época seca que se extiende de enero a marzo (Instituto Meteorológico Nacional [IMN], 2008). La temperatura media anual es de 27 ºC, la máxima de 33,5 ºC y la mínima de 22 ºC. Los suelos son inceptisoles, con alta o baja saturación de bases y entisoles con poca o excesiva humedad.
Se seleccionaron 12 plantaciones con topografía plana, suelos fértiles a moderadamente fértiles. En cada plantación se instaló una parcela de 500 m2 para la identificación de los árboles dominantes entre los más gruesos y más altos. Se cortó durante el período lluvioso un total de 23 árboles dominantes distribuidos en las edades de uno a 12 años (1; 2; 3,5; 4; 5; 5,5; 6,5; 7; 7,5; 8,5; 9 y 12 años). Cada árbol fue seccionado en cinco componentes: tronco, corteza, ramas primarias, ramas secundarias y hojas. En todas las edades se cortaron dos árboles por parcela (excepto para 8,5; solo un árbol tenía las características deseadas). Antes de talar cada árbol se limpió de hojas verdes caídas y se colocó una lona sobre la cual se dirigió la caída de este mismo. En lonas separadas extendidas sobre el suelo, se recogió la biomasa de cada una de las fracciones del árbol. Se denominó ramas primarias a aquellas que salían del tronco, mientras que las secundarias fueron aquellas que salían de una rama primaria. Se obtuvo una muestra del follaje de cada árbol, conformada por 4 grupos de 10 hojas (40 en total por árbol). La muestra de ramas secundarias se hizo con 20 trozos pequeños tomados al azar del total y se envió al laboratorio, donde fueron molidos con un triturador con cuchillas. El tronco se partió en secciones consecutivas de un metro y se procedió a quitarle la corteza. Para remover la corteza del tronco se partió longitudinalmente esta aún unida al tronco, con una motosierra y se desprendió con una herramienta en forma de espátula. Para la toma de la muestra del tronco se partieron las secciones del tronco en segmentos de 25 cm con la ayuda de una motosierra y se recogió un kg de aserrín sobre una lona limpia. La muestra de corteza y ramas primarias se obtuvo cortando con motosierra todas las secciones y recogiendo luego cerca de un kg de aserrín previamente homogeneizado para cada caso y colocado en bolsas plásticas. Antes de recoger el aserrín se aseguró la eliminación de la contaminación con basuras o cualquier pedazo de material sin cortar. En el campo, las muestras se almacenaron en bolsas de papel debidamente identificadas, se guardaron en hieleras para evitar el deterioro y se enviaron al laboratorio del Centro de Investigaciones Agronómicas de la Universidad de Costa Rica (CIA-UCR). El análisis de las muestras foliares se realizó utilizando la metodología de digestión húmeda con ácido nítrico, en el extracto se determinó el Ca, Mg, K, Zn, Cu, Mn y Fe por emisión atómica con plasma, el P por el método colorimétrico, el S por el método turbidimétrico y el N por el método Microkjeldahl (Henríquez, Bertsch y Salas, 1995). Se tomó una muestra de suelo compuesta (6 submuestras) alrededor de cada árbol cortado, en un área de 36 m2 y a 15 cm de profundidad y fueron enviadas al laboratorio del CIA de la Universidad de Costa Rica. Se determinó el pH en agua 10:25; Acidez, Al, Ca y Mg con KCl 1M 1:10; P,K,Zn,Fe,Mn y Cu con Olsen modificado pH 8,5 (NaHCO3 0,5 N, EDTA 0.01M, Superfloc 127) 1:10; S con Ca(H2PO4)2·H2O 0,008M 10:25. La acidez determinada por valoración con NaOH y Al con HCl; P y S por colorimetría con el analizador de inyección de flujo (FIA) y el resto de los elementos por espectrofotometría de absorción atómica. Se determinó el porcentaje de arena (Aren), arcilla (Arc) y limo (Limo) por el método del hidrómetro de Bouyoucos.
El análisis de los datos se basó primero en la determinación de los promedios de concentración y el intervalo de confianza (95%) de cada nutrimento en cada componente sin hacer distinción de la edad. Luego se hizo una comparación (prueba de medias por medio de la “T de student”) entre la concentración de cada nutrimento en los distintos componentes de la BA. Se realizó un análisis de correlación de la concentración entre todos los nutrimentos en cada componente. Se determinaron las curvas de mejor ajuste (tendencias) de los datos para la concentración de cada uno de los nutrimentos para cada componente, desde uno hasta los 12 años. Para determinar las tendencias se promediaron los datos de una misma edad. Luego, en otro análisis se agruparon los datos de concentración de cada nutrimento, en cada componente, en clases de edad (1 a 4; 5 a 8 y 9 a 12 años), con el fin de comparar las medias de concentración por clase etárea por medio de la “T de student”. Finalmente, se hizo una comparación entre la concentración de cada nutrimento de la BA con las variables del suelo: pH, acidez, SA y CICE. Todos aquellos coeficientes de correlación mayores de 0,59 se consideraron como correlaciones aceptables. Los datos se analizaron con el programa Estadística StatSoft, Inc. (2001) STATISTICA (Data Analysis Software System), versión seis.
3. Resultados y discusión
Los componentes de la BA con la mayor concentración total de macronutrimentos fueron las hojas (6,3%), seguidos de la corteza (3,57%), las ramas secundarias (2,56%), las ramas primarias (1,72%) y el fuste (0,78%) (Cuadro 1). Los macroelementos de mayor concentración en todos los componentes fueron el N, Ca y K y los menos abundantes fueron el Mg, P y S. En todos los componentes el orden de mayor a menor concentración del Mg, P y S se mantuvo invariable, sin embargo, el orden del N, Ca y K varió según el componente, de tal modo que el N fue el macroelemento más concentrado en las hojas, pero el Ca es más concentrado en la corteza y las ramas primarias, no obstante, el K es más concentrado en el fuste y las ramas secundarias. Similar información fue obtenida por Negi et al. (1990).
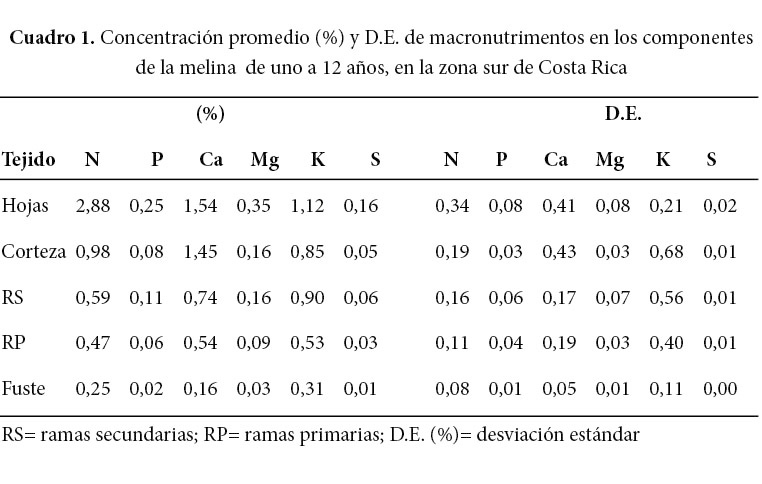
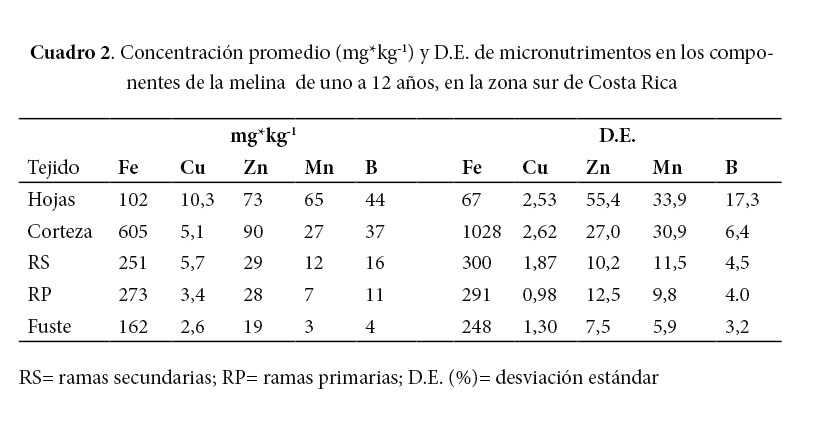
El componente de la BA con la mayor concentración de micronutrimentos fue la corteza (765 mg kg-1), seguido de las ramas primarias (323 mg kg-1), las ramas secundarias (314 mg kg-1), las hojas (295 mg kg-1) y el fuste (190 mg kg-1). El Fe, el Zn y el B fueron los micronutrimentos más abundantes en todos los componentes de la BA, excepto en las hojas, en las cuales el orden fue Fe, Zn y Mn. El Mn y el Cu fueron los micronutrimentos menos abundantes en todos los componentes de la BA, excepto en las hojas. El valor promedio tan alto del hierro puede ser evidencia de contaminación a causa de partículas de suelo en el aire.
La concentración foliar de todos los nutrimentos de la BA (Cuadro 1 y Cuadro 2) es similar a la reportada por Avellán (2012), excepto para el Zn, cuyo valor es 30% superior al reportado por ese autor. Avellán (2012) estudió la variación de la concentración foliar según la época y encontró que la concentración foliar de N, K, Fe y Al disminuyó en la época seca y la concentración de Ca y Mn aumentó en la época seca. Avellán (2012) también hizo el análisis de la concentración foliar según la copa superior y la copa media de los árboles de melina y encontró una disminución de 15% (no significativa) en la concentración de Ca en la copa superior con respecto a la copa media, probablemente debido a la poca movilidad de este elemento y encontró que la concentración de Ca y Zn disminuye en las hojas externas de las ramas, en comparación con las internas.
Al comparar los datos de esta investigación con los de Negi et al. (1990) en la India para árboles de 20 años, se nota que las concentraciones de macronutrimentos en las ramas primarias y secundarias son similares, pero hay diferencias en las hojas, la corteza y el fuste, ya que los valores de N y K de las hojas y la corteza son mayores en el presente estudio, lo mismo que el Ca en el fuste, mientras que la concentración de Ca foliar fue mayor en la India (Negi et al., 1990).
Arias et al. (2011) encontraron en árboles de seis años contenidos menores para la concentración del P en todos los componentes de la BA, Ca en las hojas y en la corteza, N en la corteza y el tallo, K en las hojas y el tallo, Mg en la corteza, en comparación con este estudio. Arias et al. (2011) encontraron valores superiores en la concentración del N, K, Ca y Mg en las ramas, Ca en el tallo y Mg en las hojas y en las ramas, con respecto a los de esta investigación.
Se determinaron los intervalos de confianza de la concentración de cada uno de los nutrimentos en los distintos componentes de la BA (Cuadro 3).
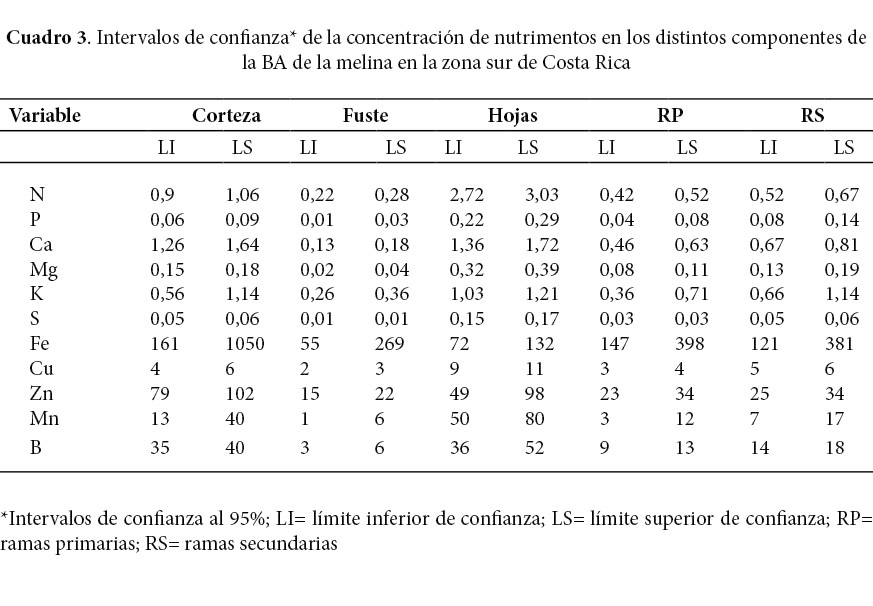
Los intervalos de concentración foliar determinados para el K, N, Mg, P, S, Fe, Mn, B y Cu son semejantes a los de Avellán (2012), sin embargo, los intervalos de concentración foliar del Ca y el Zn fueron mayores que los reportados por el mismo autor.
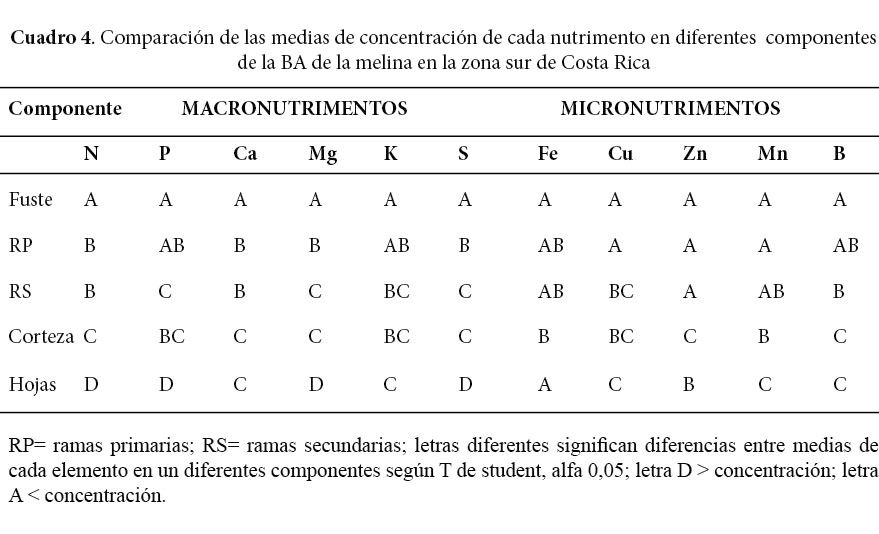
Se encontraron diferencias altamente significativas entre las medias de concentración de los macronutrimentos (N; P; Ca; Mg; K y S) y los micronutrimentos (Cu; Zn; Mn; y B) en los distintos componentes, sin embargo, no se determinaron diferencias significativas entre las medias de Fe en los distintos componentes (Cuadro 4).
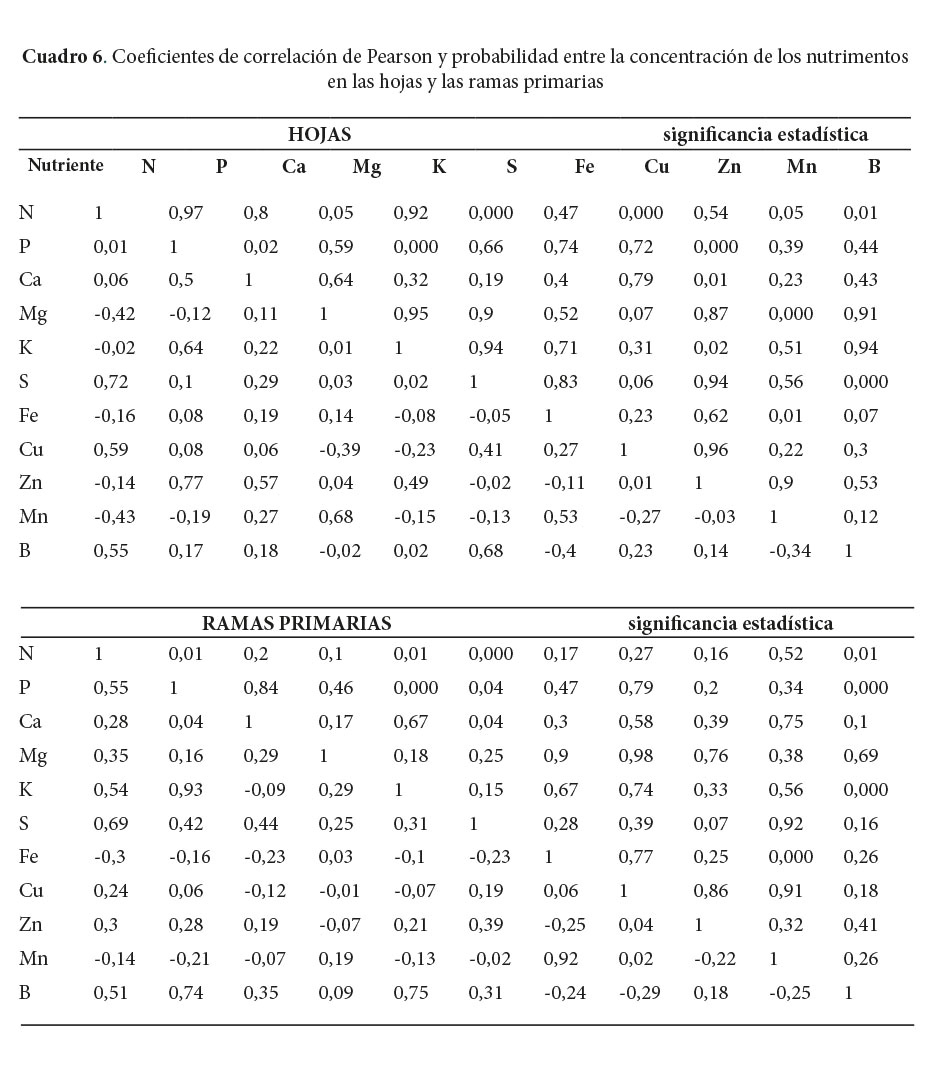
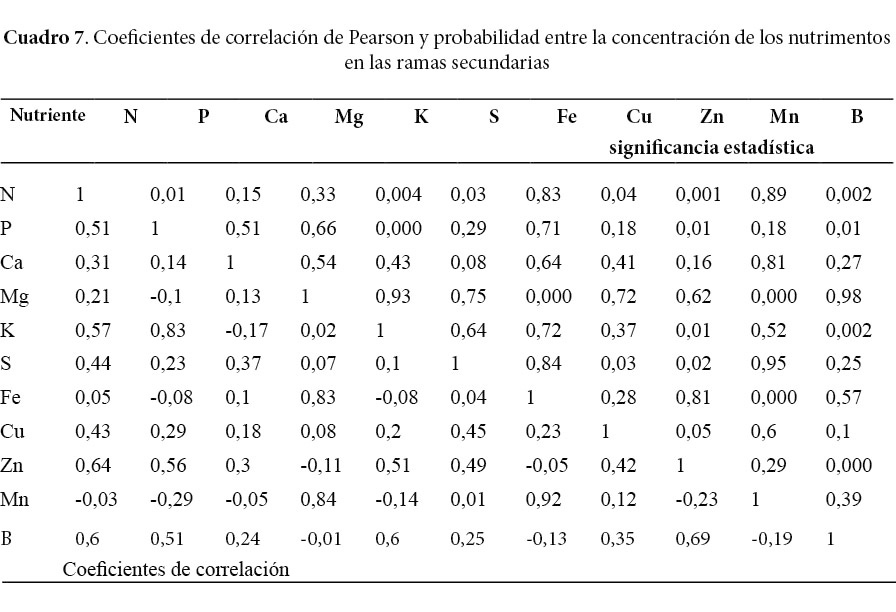
Todas las correlaciones de Pearson entre 0,5 a uno fueron positivas y significativas, mientras que las correlaciones negativas mantienen correlaciones calificadas como bajas (Cuadros 5, 6 y 7). El Fe y el Mn tienen correlaciones positivas en todos los componentes de la biomasa y además, fueron los nutrimentos con las correlaciones más altas encontradas en este estudio. El P mantiene correlaciones positivas mayores de 0,5 con el K; Mg; Zn; B y el N; además se destaca la fuerte relación del P con el K en el fuste, la corteza y las ramas primarias. El N mostró una buena correlación con el S de la corteza y las hojas y el P del fuste.
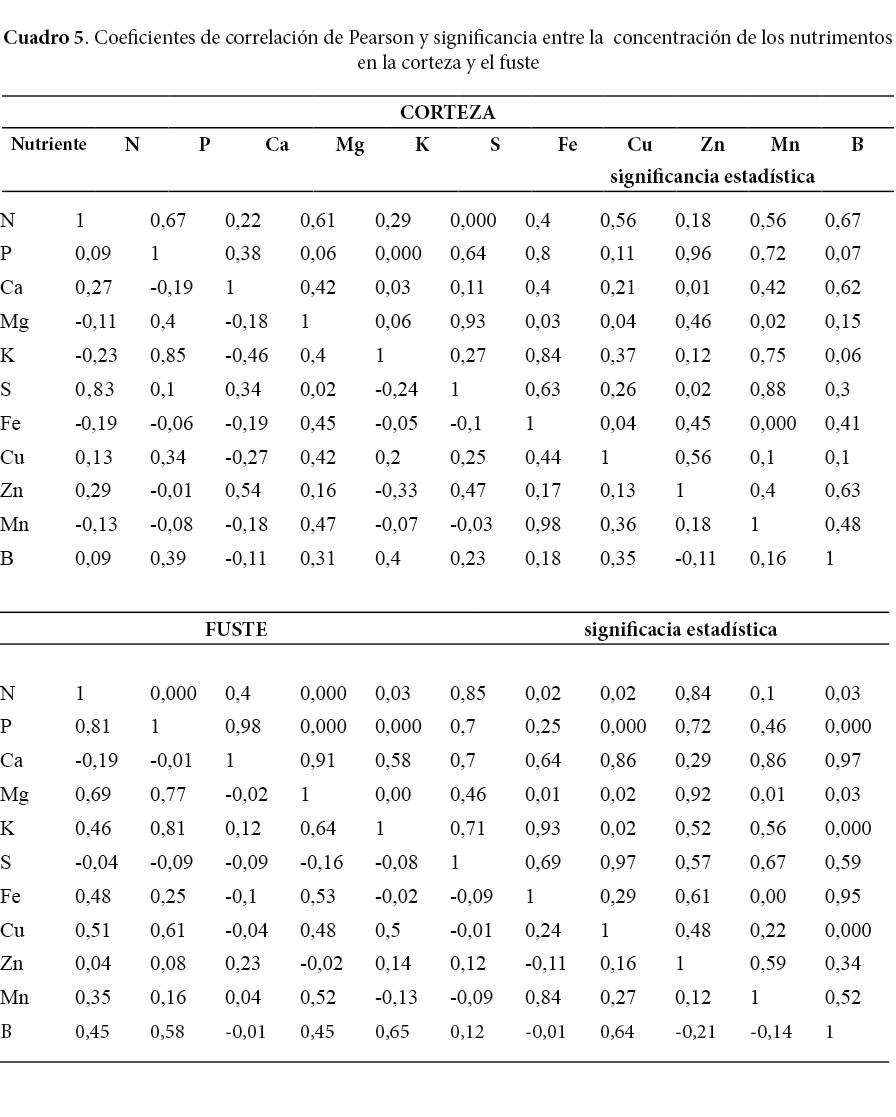
Las correlaciones negativas, aunque menores de 0,5, se presentaron entre el K con el N; Ca; Cu y Zn en la corteza (Cuadro 5). Por otro lado, se evidenciaron correlaciones negativas entre el N con el Mg y el Mn en las hojas, además del Cu con el Mg y el K en el mismo componente (Cuadro 6). En el caso de correlaciones negativas entre nutrimentos de los componentes ramas primarias y secundarias, se encontró básicamente entre micronutrimentos como, por ejemplo, el Mn con el Zn y el B de ambos componentes (Cuadros 6 y 7).
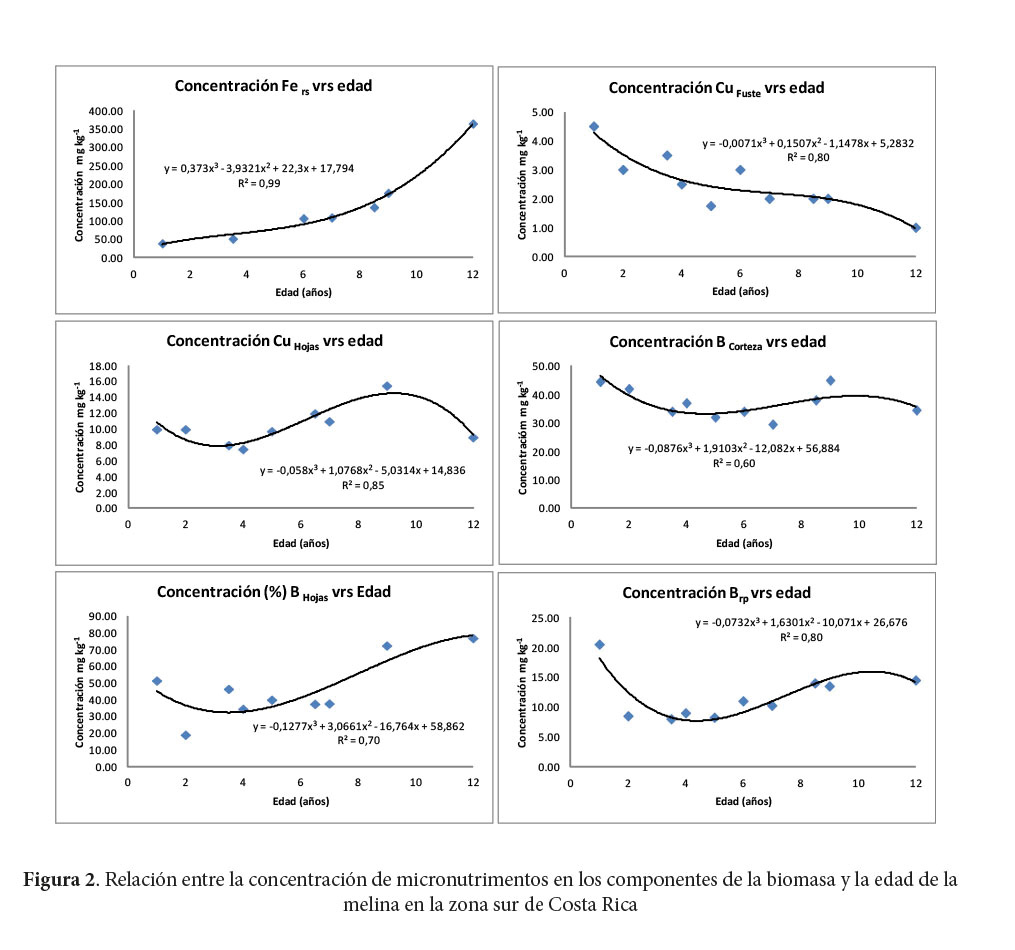
Los macronutrimentos cuya concentración se puede atribuir a la diferencia de la edad de la plantación en más de 90% (R2) fueron el N, P y Mg en el fuste y el Ca en las hojas (Figura 1). El Ca es el macronutrimento que es explicado en más de 70% por la edad de la plantación en cuatro de los componentes de la BA (la corteza, las hojas, las ramas primarias y secundarias). La concentración de K en el fuste, las hojas y las ramas secundarias, también se explica satisfactoriamente (> 70%) por la edad de la plantación. El fuste es el componente de la BA que mantiene buena relación de explicación de la concentración de N, P, K y Mg, además, la corteza es el componente que explica la concentración de Ca, P, K y Mg según la edad. Se encontró que la concentración de N, Ca y K en las hojas puede ser explicada satisfactoriamente por la edad.
La concentración de micronutrimentos que mayor relación tiene con la edad de la plantación de melina es la del Fe (R2= 0,99) en las ramas secundarias. El único microelemento cuya concentración puede ser explicada por la edad de los árboles en varios componentes de la BA, es el B en las hojas, la corteza y las ramas primarias.
El Ca es el único macroelemento con tendencia a aumentar su concentración con la edad de los árboles en todos los componentes de la BA. Avellán (2012) encontró que el Ca foliar de árboles de nueve a 12 años fue 26% superior que en árboles más jóvenes. Onweremadu (2007) determinó un aumento de la concentración foliar de Ca con la edad en plantaciones de melina, a pesar de una disminución de su contenido en el suelo con el tiempo.
Con el aumento en edad de la plantación, el NHojas tiene la tendencia a aumentar la concentración, mientras que, el NFuste tiene la tendencia a disminuir con el aumento en edad.
El K es el único macroelemento en todos los componentes que tiene la tendencia a disminuir la concentración con el aumento en edad de los árboles. Lo mismo fue hallado por Avellán (2012), quién determinó que el K foliar de árboles de uno a cuatro años disminuyó en 25% respecto de árboles de nueve a 12 años. Otros macroelementos con tendencia a disminuir concentración en alguno de los componentes fueron: NFuste; PCorteza; PFuste y el MgCorteza y MgFuste.
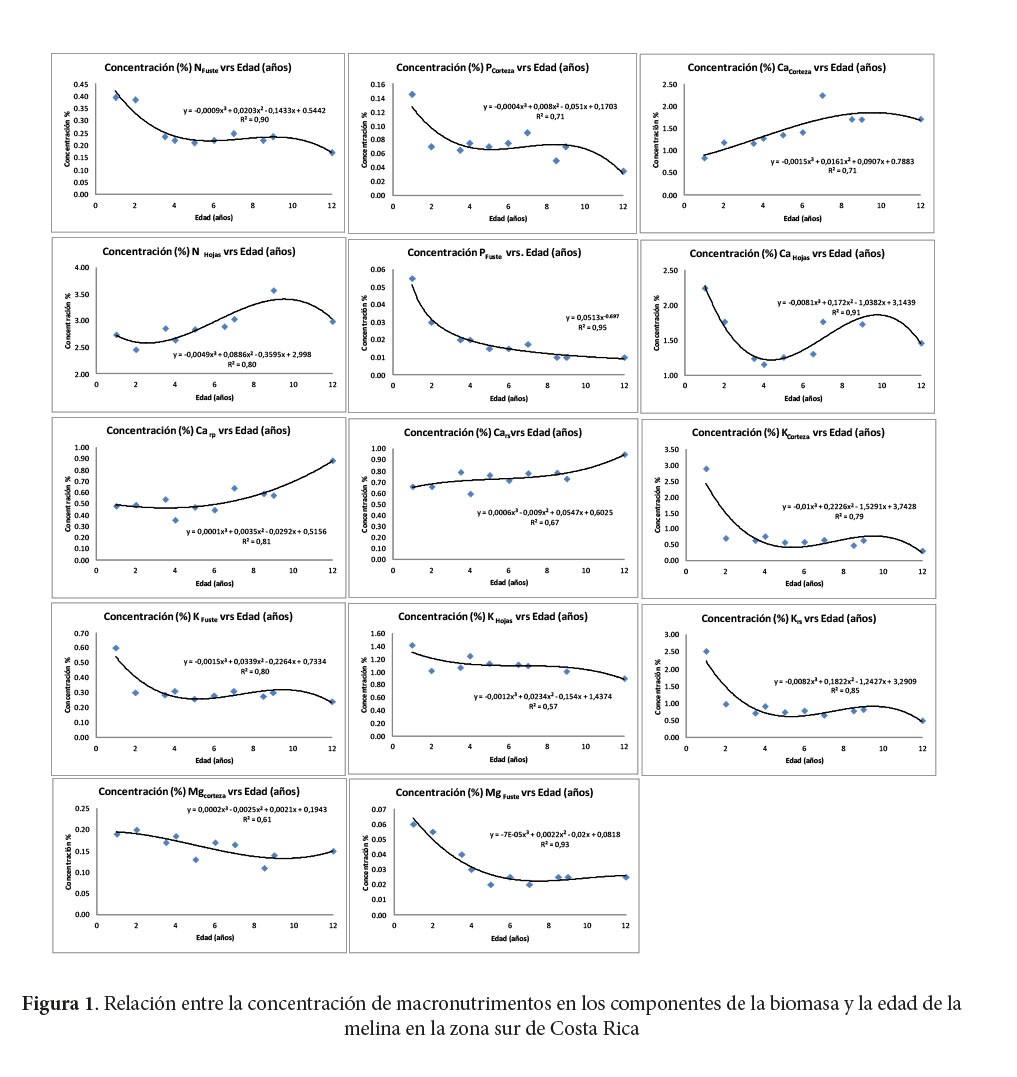
Los micronutrimentos que tienen una tendencia a aumentar su concentración con la edad fueron el FeRamas Secundarias, BHojas y el CuHojas que primero aumenta y luego disminuye. Según Avellán (2012), la concentración foliar de B aumentó 55% desde edades jóvenes (uno a cuatro años) a edades maduras (nueve a 12 años). Los micronutrimentos que tienen la tendencia a disminuir su concentración con la edad fueron el CuHojas y el BCorteza y el BRamas primarias, que merma primero y luego aumenta.
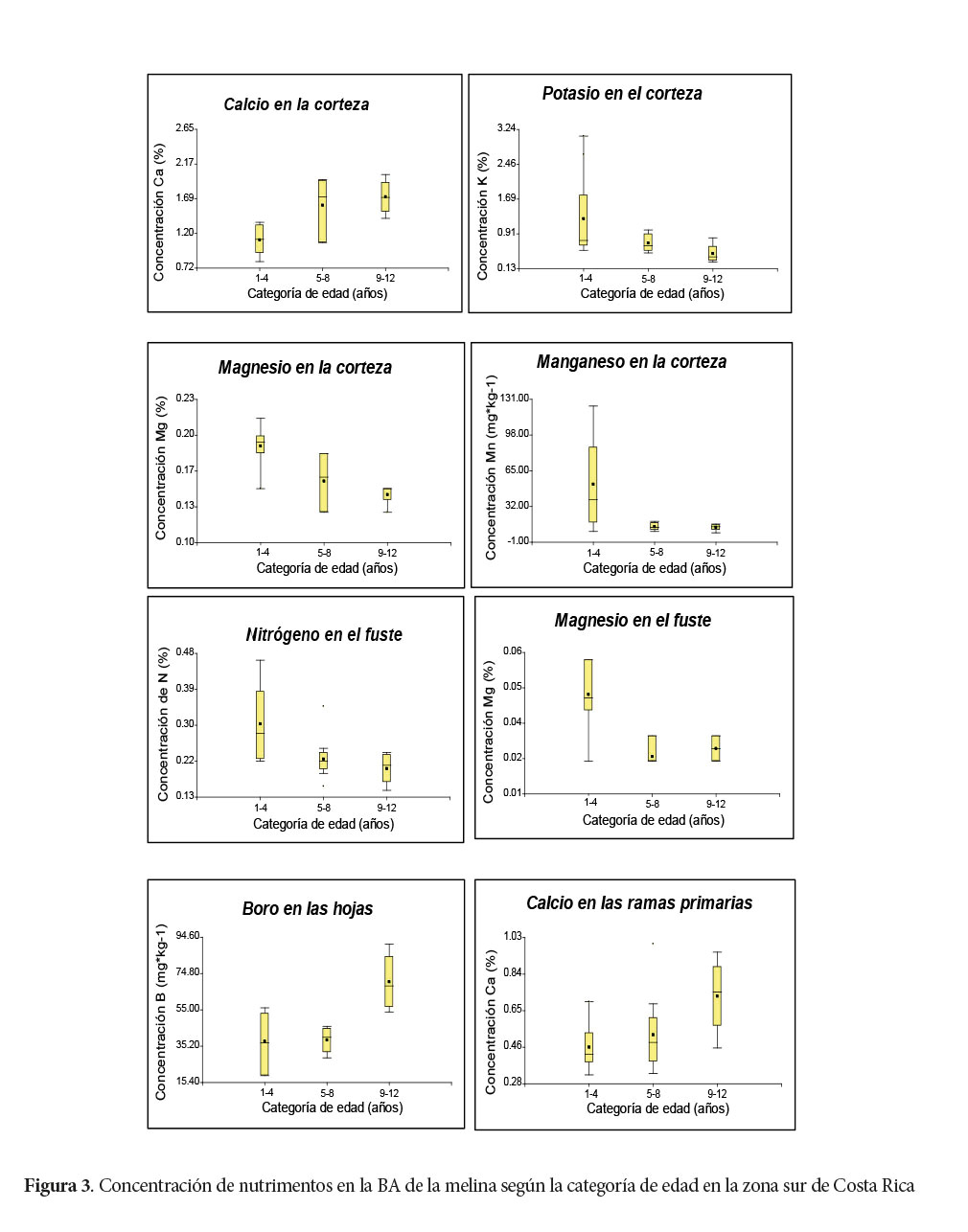
Se registraron diferencias significativas al 95% entre los promedios de concentración de las categorías de edad 1-4 y 9-12 años para los nutrimentos Ca; Mg y Mn en la corteza, Mg en el fuste, B en las hojas y Ca en las ramas primarias. En la concentración de K en la corteza, aunque tiene una tendencia a disminuir con el aumento en las categorías de edad, las diferencias no fueron significativas.
3.1 Relación entre algunas propiedades del suelo y la concentración de nutrimentos en los BA de melina
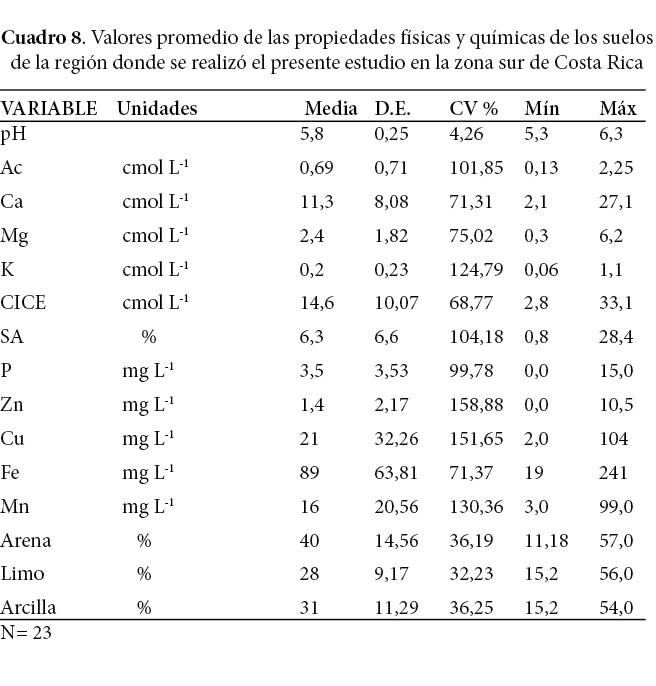
Los suelos (inceptisoles y entisoles aluviales) de la región donde se llevó a cabo el estudio pueden considerarse como adecuados para el buen desarrollo de plantaciones de melina, según los criterios desarrollados por Murillo y Alvarado (2012), es decir, sin deficiencias en el contenido de bases y un rango de saturación de acidez entre 0,81% y 28% (Cuadro 8). Son suelos planos o de poca pendiente, con pH entre 5,3 y 6,3. El pH es la variable que tiene menor dispersión, todas las demás presentan coeficientes de variación entre 32 y 158%, lo que se debe a la presencia de suelos inceptisoles de bajos y altos contenidos de bases y a entisoles de altos contenidos de bases. El contenido de bases en los suelos estudiados tiene una correlación de 0.78 con el contenido de arcilla (%) y una correlación negativa de -0.75 con el contenido de arena en el suelo.
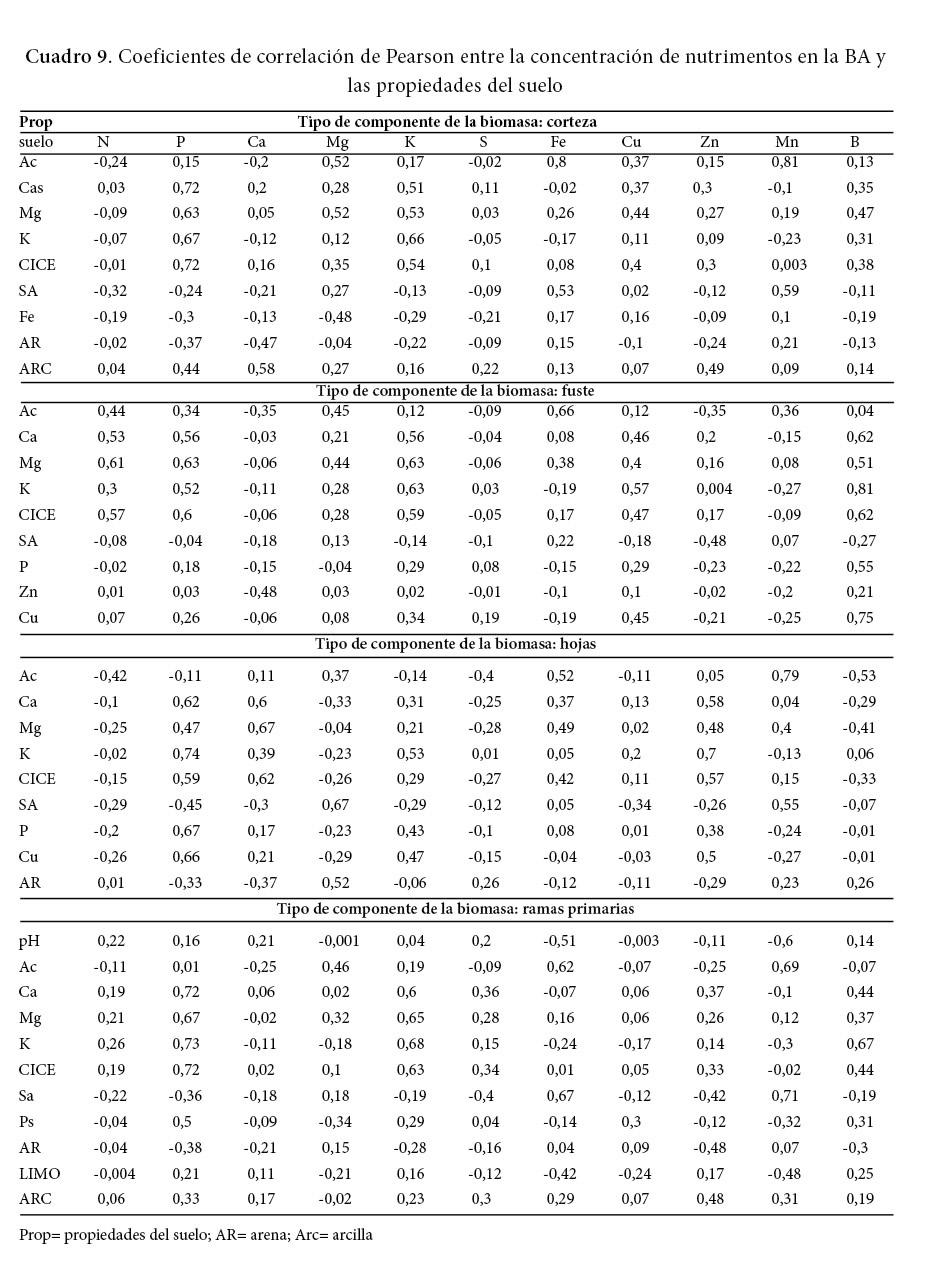
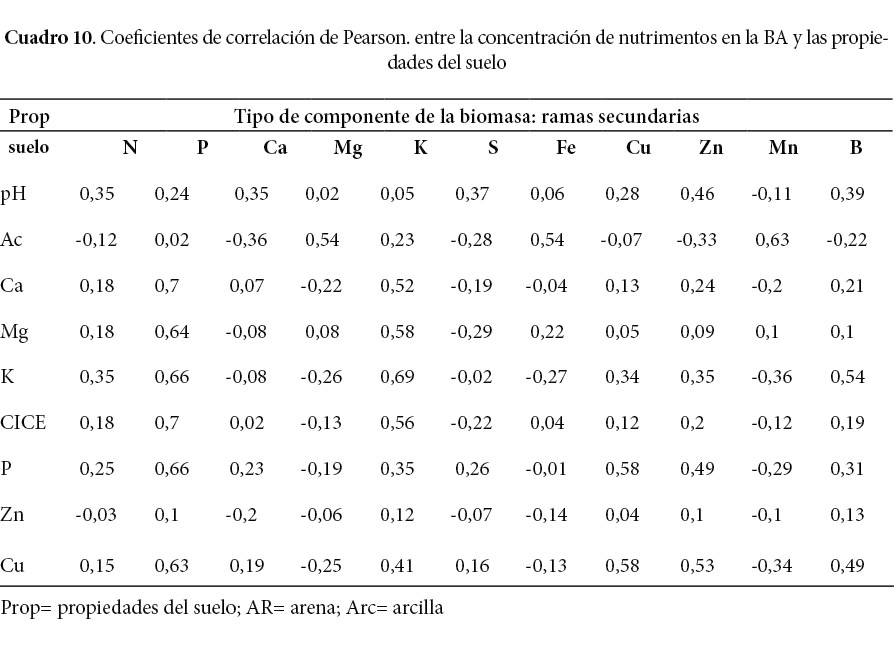
Las correlaciones más fuertes encontradas (Cuadro 9 y 10) fueron entre la concentración de Fe y Mn (coeficientes de 0,8 y 0,81 respectivamente) en la corteza de los árboles con la acidez del suelo. Además, existe una fuerte correlación entre la concentración de B en el fuste y el K (coeficiente de 0,8) del suelo (Cuadro 9). En todos los componentes de la BA hay una correlación positiva media a alta de la concentración de Fe y Mn con la acidez extraíble del suelo (Ac), lo que significa que, al aumentar la acidez del suelo, podría aumentar la disponibilidad de estos elementos en el suelo.
El análisis de las correlaciones entre cada nutrimento en el suelo y su concentración en los componentes de la biomasa reflejó que el K del suelo tiene correlaciones altas (0,66; 0,63; 0,68; 0,69) con la concentración de este nutrimento en la corteza, el fuste y las ramas primarias y secundarias respectivamente, además mantiene una correlación media (0,53) con la concentración en las hojas. Lo anterior indica que la melina responde a los aumentos de K en el suelo y lo absorberá. Escobar (2013) encontró una correlación un poco más baja (0,44) entre el K foliar y el K disponible del suelo, sin embargo, este autor realizó su estudio de la relación de los nutrimentos disponibles en el suelo y la concentración foliar en la melina basándose mayormente en suelos vertisoles y algunos inceptisoles. Escobar (2013) halló, también, una correlación fuerte (r =0,68) entre la concentración de sodio de la fase intercambiable y la concentración de sodio de la fase soluble y, además, halló que el crecimiento de la melina fue mejor en sitios con sodio intercambiable de menor concentración, no obstante, ni Escobar (2013) ni Caguasango (2012) encontraron una relación aceptable entre el sodio foliar y el sodio de la solución o el intercambiable del suelo. En la presente investigación no se analizó el sodio porque son suelos con bajos contenidos y poca variabilidad (Murillo, 1996), pero, podría ser un factor de importancia en suelos básicos (terrenos planos o de poca pendiente en el noroeste) de otras zonas de Costa Rica como Guanacaste. Caguasango (2012) registró una relación negativa baja del sodio de la solución del suelo con el P (r = -0.31) y el Ca foliar (r = -0,29) de la melina.
El P es otro elemento con correlaciones medias a altas entre su contenido en el suelo y la concentración en las hojas, las ramas primarias y secundarias.
Osundina y Osonubi (1989) hallaron que hubo una disminución de la concentración de los elementos N, K y Mn y un aumento de P y Fe en el suelo, causada por la inundación del suelo, lo que provocó igual comportamiento en la concentración foliar de estos elementos. Onwermadu (2007) encontró una disminución del contenido de Ca en el suelo; sin embargo, la concentración foliar aumentó.
4. Conclusiones
Los resultados de este estudio son una base para el diagnóstico nutricional y para elaborar programas de fertilización con melina, especialmente en la zona sur del país y aquellas similares en suelos y régimen de precipitación. La mayor concentración de macronutrimentos se encontró en las hojas y la de micronutrimentos en la corteza, mientras que la menor concentración de macro y micronutrimentos se mostró en el fuste. Los macroelementos de mayor concentración en todos los componentes fueron el N, Ca y K y los menos abundantes fueron el Mg, P y S. El Fe, el Zn y el B resultaron los micronutrimentos más abundantes en todos los componentes excepto en las hojas. Se encontró diferencias altamente significativas entre las medias de concentración de los macronutrimentos (N; P; Ca; Mg; K y S) y los micronutrimentos (Cu; Zn; Mn; y B) en los distintos componentes; pero, no se evidenciaron diferencias estadísticas entre las medias de Fe en los distintos componentes.
La concentración de Ca tiene la tendencia a aumentar con la edad de los árboles en todos los componentes de la BA, mientras que el K posee la tendencia a disminuir. El NHojas la tendencia es a aumentar la concentración con la edad; sin embargo, en el NFuste es a disminuir con el aumento en edad.
Existe una fuerte correlación (coeficientes de 0,8 y 0,81) entre la concentración de Fe y Mn (respectivamente) en la corteza de los árboles, con la acidez del suelo. También, se encontró que el K del suelo tiene correlaciones medias a altas (0,66; 0,63; 0,68; 0,69) con la concentración de este nutrimento en la corteza, el fuste y las ramas primarias y secundarias. Asimismo, la concentración de Ca y Mg en el suelo está bien correlacionado (0,6 y 0,67 respectivamente) con la concentración de esos elementos en las hojas.
5. Agradecimientos
Al personal de la Revista de Ciencias Ambientales y a las personas revisoras, quienes con sus aportes mejoraron el artículo.
6. Bibliografía
Agus, C., Karyanto, O., Hardiwinoto, S., Haibara, K. y Toda, H. (2000a). Biomass in fast growing species of tropical Gmelina arborea Roxb. plantation forest. In Transaction of the 111st Japanese Forestry Society (pp.557-558).
Agus, C., Sumardi, O., Hardiwinoto, S., Na’iem, M., Sipayung, M., Wardana, W., Haibara, K. y Toda, H. (2000b). Biomass of carbon and nutrient stock in Gmelina arborea Roxb. plantation forest. Proceeding Seminar on Forest Biomass. GMU-JIFPRO. Yogyakarta.
Agus, C., Karyanto, O., Kita, S., Haibara, K., Toda, H., Hardiwinoto, S., Supriyo, H., Naiem, M., Wardana, W., Sipayung, M.S., Wijoyo, K. y Wijoyo, S. (2004). Sustainable site productivity and nutrient management in a short rotation plantation of Gmelina arborea in East Kalimantan, Indonesia. New Forest, (28), 277–285. doi: http://dx.doi.org/10.1023/B:NEFO.0000040954.27630.2f
Arce, H. (2012). Plantaciones forestales establecidas mediante pago de servicios ambientales (PSA). En presentación sobre “Informe Interno Anual FONAFIFO 2012”. Sp.
Arias, D., Calvo, A., Richter, D. de B. y Dohrenbusch, A. (2011). Productivity, aboveground biomass, nutrient uptake and carbon content in fast-growing tree plantations of native and introduced species in the Southern Region of Costa Rica. Biomass and bioenergy, 35, 1779-1788. doi: http://dx.doi.org/10.1016/j.biombioe.2011.01.009
Avellán, M. J. (2012). Determinación y variación de la concentración foliar de nutrimentos de Gmelina arborea Roxb. en Osa, Corredores y Golfito, Costa Rica (Tesis de Licenciatura). Universidad Nacional de Costa Rica,
Caguasango, S. M. (2012). Modelación de la calidad de índice de sitio – suelo sitio específico mediante criterios de disponibilidad de nutrientes en plantaciones de Gmelina arborea y Pachira quinata en bosque seco tropical (Zambrano – Bolívar) (Tesis). Universidad Distrital Francisco José de Caldas Bogotá, Colombia.
Correndo, A. y García, F. (Marzo, 2012). Concentración de nutrimentos en planta como herramienta de diagnóstico: Cultivos extensivos. Archivo Agronómico #14, IPNI International Plant Nutrition Institute.
Drechsel, P. y Zech, W. (1991). Foliar nutrient levels of broad-leaved tropical trees: A tabular review. Plant and Soil, 131(1), 29-46.
Escobar, L. J. (2013). Relación de parámetros de fertilidad del suelo con el índice de sitio determinado para plantaciones forestales de melina (Gmelina arbórea) y ceiba (Pachira quinata) en Zambrano-Bolivar (Colombia) (Tesis de maestría). Universidad Nacional de Colombia.
Evans, J. (1979). The effects of leaf position and leaf age in foliar analysis of Gmelina arborea. Plant and Soil, 52(4), 547-552. doi: http://dx.doi.org/10.1007/BF02277950
Henríquez C., Bertsch, F., Salas R. (1995). Fertilidad de suelo: Manual de laboratorio. San José, Costa Rica: Asociación Costarricense de la Ciencia del Suelo.
Holdridge, L. (1982). Ecología basada en zona de vida (Trad. del inglés por Jiménez, H. Segunda reimpresión). Instituto Interamericano de Cooperación para la Agricultura. San José, Costa Rica.
Instituto Meteorológico Nacional (IMN). (2008). Clima, variabilidad, cambio climático en Costa Rica. Recuperado de http://www.slideshare.net/Socialesdigital/el-clima-variabilidad-y-cambio-climatico-en-cr
Kok, R. (2012). A revision of the genus Gmelina (Lamiaceae). Kew Bulletin, (67), 293-329.
Murillo, R. (1996). Evaluación de algunos factores ambientales que afectan la calidad de sitio a nivel de micrositio para melina (Gmelina arborea Robx) plantada en suelos planos en la zona Sur de Costa Rica (Tesis de licenciatura). Universidad Nacional. Costa Rica.
Murillo, R. y Alvarado, A. (2012). Nutrición y fertilización de Gmelina arborea. En A. Alvarado y J. Raigosa (Eds.), Nutrición y fertilización forestal en regiones tropicales (pp. 285-299, 1era ed.). San José, Costa Rica: Asociación Costarricense de la Ciencia del Suelo.
Negi, J. D. S., Bahuguna, V. K. y Sharma, D. C. (1990). Biomass production and distribution of nutrients in 20 years old teak (Tectona grandis) and Gamar (Gmelina arborea) plantation in Tripura. Indian Forester, 116(9), 681-686.
Ogbonnaya, C. y Kinako, P. (1993). Growth and mineral nutrition of Gmelina arborea Roxb. Tree Physiology, 12, 291-299.
Olaleye, A. O., Aderinola, A. P., Onasanya, A. S. y Akinbola, G. E. (2002). Utilization of Gmelina aborea (Roxb) to ameliorate a degraded Alfisol in South-Western Nigeria.
Onweremadu, E. U. (2007). Pedogenic loss and uptake of calcium by Gmelina growing in na Isohyperthermic Kandiudult. Journal of Plant Sciences, 2(6), 625-629. doi: http://dx.doi.org/10.3923/jps.2007.625.629
Onyekwelu, J., Mosandl, R. y Stimm, B. (2006). Productivity, site evaluation and state of nutrition of Gmelina arborea plantations in Oluwa and Omo forest reserves, Nigeria. Forest Ecology and Management, 229, 214–227. doi: http://dx.doi.org/10.1016/j.foreco.2006.04.002
Osundina, M. A. y Osonubi, O. (1989). Adventitious roots, leaf abscission and nutrient status of flooded Gmelina and Tectona seedlings. Tree Physiology, 5, 473-483. doi: http://dx.doi.org/10.1093/treephys/5.4.473
Sampaio, LM. y Huber, W.E. (1999). Concentração e distribuição de nutrimentos nas folhas de espécies florestais, na Amazônia Ocidental. Floresta e Ambiente, 6(1), 127-137.
Stuhrmann, M., Bergmann, C. y Zech, W. (1994). Mineral nutrition, soil factors and growth rates of Gmelina arborea plantations in the humid lowlands of northern Costa Rica. Forest Ecology and Management, 70, 135-145. doi: http://dx.doi.org/10.1016/0378-1127(94)90081-7
