Rev. Mar. Cost. ISSN 1659-455X. Vol. 3: 171-181, Diciembre 2011.
DOI: https://doi.org/10.15359/revmar.3.13
ESTUDIO DE LAS POBLACIONES DE CABALLITOS
DE MAR EN DOS ZONAS DE LA COSTA NORTE
DE LA HABANA Y PINAR DEL RÍO, CUBA
STUDY OF SEAHORSE POPULATIONS IN TWO AREAS
IN THE NORTH COAST OF LA HAVANA AND
PINAR DEL RÍO, CUBA
Lourdes Pastor Gutiérrez1*, Yuliet Piloto Cubero1, Raúl Igor Corrada Wong1 y Pedro Pablo Chevalier Monteagudo1
1 Acuario Nacional de Cuba. Ministerio de Ciencia, Tecnología y Medio Ambiente. Ave. 1ra. No. 6024. Miramar. Playa. La Habana CP 11300 Cuba. *lourdesp@acuarionacional.cu
Recibido 24-IV-2011
Aceptado 09-IX-2011
RESUMEN
Se presentan los resultados sobre el estudio de las poblaciones de caballitos de mar en dos estaciones ubicadas en la costa norte de La Habana y Pinar del Río, Cuba. Se llevaron a cabo censos visuales mediante buceo libre, utilizando el método de transectos lineales, entre los meses de abril del 2004 y junio del 2005. Se obtuvieron la densidad media para las especies Hippocampus reidi (Ginsburg, 1933) e Hippocampus erectus (Perry, 1810), así como la densidad media por estaciones y por época del año. La especie H. reidi fue la más abundante y la mayor densidad se encontró en la estación de Las Cuarenta 0.0109 ind/m2. No se encontraron diferencias entre la densidad en época de lluvia y seca para las especies. Los machos y las hembras de H. reidi mostraron tallas similares (LT). Se supone un comportamiento monógamo de la especie H. reidi, debido a la proporción sexual encontrada, muy similar a la proporción esperada de 1:1.
Palabras claves: Hippocampus reidi, Hippocampus erectus, caballito de mar, densidad, Cuba.
ABSTRACT
This paper presents the results of the study on seahorse populations at two stations located in the north coast of La Havana and Pinar del Río, Cuba. Visual census were conducted by free divers, using the linear transect method between April 2004 and June 2005. Mean density was obtained for the species Hippocampus reidi (Ginsburg, 1933) and Hippocampus erectus (Perry, 1810), by stations and by seasons of the year. H. reidi was the most abundant species and the biggest density was present in Las Cuarenta station 0.0109 ind/m2. No differences were found regarding density between the rainy and dry seasons for the studied species. Male and female H. reidi showed similar sizes (TL). A monogamous behavior of the species H. reidi is inferred due to the sexual proportion found, which is very similar to the expected proportion of 1:1.
Keywords: Hippocampus reidi, Hippocampus erectus, seahorse, density, Cuba.
INTRODUCCIÓN
En el mundo, los caballitos de mar son unas de las especies marinas más afectadas por la sobreexplotación y la degradación del ambiente (Vincent, 1996; Foster & Vincent, 2004), siendo un objetivo muy importante para los estudios y planes de conservación de la diversidad marina. Por esta razón, desde el año 2004, todas las especies del género Hippocampus están incluidas en el APÉNDICE II de la Convención Internacional sobre el Comercio Internacional de Especies Amenazadas de Flora y Fauna Silvestres (CITES, 2004), y en la Lista Roja de especies amenazadas (UICN, 2004).
Hasta el año 2004 aparecían descritas 33 especies dentro del género Hippocampus (Lourie et al. 2004). En la actualidad se han encontrado cinco nuevas especies, según Lourie & Kuiter (2008). En Cuba solamente se han registrado tres de estas: H. erectus; H. reidi e Hippocampus zosterae Jordan y Gilbert, 1882. En Cuba, la exportación de caballitos de mar vivos de las especies Hippocampus reidi Ginsburg, 1933 y de Hippocampus erectus Perry, 1810, se realizó entre los años 1995 y 2005 (UNEP-WCMC CITES, 2006), destacándose el año 2004 con más de 100 individuos vivos de la especie H. erectus. Actualmente sus poblaciones se encuentran sometidas a una presión de explotación, si consideramos que la pesca furtiva con diferentes fines (por ejemplo, acuariofilia, ornamental y medicina tradicional) es una práctica común.
La especie H. erectus se encuentra en la categoría de Vulnerable (VU) en la LISTA ROJA de la UICN (2006), debido a que sus poblaciones han disminuido al menos en un 30% en el mundo por ser una de las más comercializadas (UICN, 2009), mientras que H. reidi e H. zosterae están en la categoría taxón con Datos Deficientes (DD), por no contar con información suficiente sobre su distribución y estatus poblacional en el mundo y, por lo tanto, el riesgo real en que se encuentran no puede ser valorado actualmente. Esto conduce a que se realicen investigaciones sobre abundancia y distribución, que puedan mostrar cuál es la clasificación más apropiada para estas especies (Project Seahorse, 2003).
En general son pocos los estudios que se han realizado sobre las poblaciones naturales de caballitos de mar. Entre los trabajos más importantes sobre la especie H. reidi se encuentran los realizados por Dias & Rosa (2003); Rosa et al. (2005; 2007); Freret-Meurer & Andreata (2008); Mai & Rosa (2009) en Brasil, Arcos-Pulido & Gómez-Prieto (2004) en Santa Marta, Colombia. Teixeira y Musick (2001) estudiaron los hábitos alimenticios y reproductivos de la especie H. erectus en la bahía de Chesapeake, Estados Unidos, y Baum et al. (2003) determinaron aspectos biológicos de individuos de esta misma especie capturados por pesca incidental en el Golfo de México.
Este trabajo tiene como objetivos realizar una primera evaluación sobre el estado de las poblaciones de caballitos de mar en dos zonas de la costa norte de Cuba, de las que tradicionalmente se han extraído individuos para la exhibición en el Acuario Nacional de Cuba, así como conocer aspectos importantes sobre la abundancia, tamaño del grupo, proporción sexual y época de reproducción.
MATERIALES Y MÉTODOS
Descripción de la zona de estudio
Se seleccionaron para el estudio dos estaciones en la costa norte de Cuba (Fig. 1): Las Cuarenta ubicada entre los 22° 58. 33’ N y 82° 57. 52’ W, en la Ensenada de Silvera, Bahía de Cabañas, provincia de Pinar del Río, y El Casino entre los 23° 04. 96’ N y los 82° 30. 42’ W dentro de la Laguna de Santa Fe, provincia de Ciudad de La Habana.
Las Cuarenta: Esta zona se caracteriza por la presencia de costas bajas rocosas interrumpidas por playas con fondos cubiertos de abundante vegetación Thalassia testudinium Banks ex Koning, 1805 y una amplia extensión de mangle rojo Rhizophora mangle L. que bordea toda la zona interior (Fig. 1), presentando un fondo fangoso fundamentalmente.
Dentro de las raíces de mangle se refugian abundantes especies marinas como peces e invertebrados típicos de este ecosistema; entre los que se encuentran, esponjas: Amphimedon viridis (Duchassaing y Michelotti, 1864), Suberites aurantiaca (Duchassaing y Michelotti, 1864) y Dysidea etheria (Laubenfels, 1936), crustáceos, bivalvos, poliquetos tubícolas, ascidias, juveniles de peces, entre otros organismos. También crecen varias especies de algas como: Acetabularia crenulata J.V. Lamouroux y Bostrychia tenella (J. V. Lamouroux) J. Agardh. Frente al manglar se ubica un extenso pastizal en el que habitan equinodermos, moluscos, crustáceos y diferentes especies de algas como: Penicillum sp., Udotea sp., Rhipocephalus sp. y Caulerpa sp.
El Casino: Este sitio se encuentra dentro de un canal artificial de más de 100 m de longitud (Fig. 1), presenta dos muros grandes con una parte sumergida y un fondo de arena y piedra sueltas con crecimiento de algas. Las paredes que están debajo del agua se encuentran cubiertas por algas (Caulerpa sertularioides (S. Gmelir) M. Howe, 1905; Halimeda sp.), esponjas, ascidias, poliquetos tubícolas, erizos, entre otros. En el extremo del canal que da hacia el mar hay un pequeño parche de T. testudinum.
Método de muestreo
Para el estudio de la densidad poblacional, se llevaron a cabo censos visuales durante el horario de la mañana entre las 8 y las 10 a.m., mediante buceo en apnea, entre los meses de abril del 2004 y junio del 2005, a una profundidad que osciló entre 1 y 2 m. En Las Cuarenta se realizaron 4 muestreos en época de lluvia y 4 en la seca, y en cada uno se hicieron 16 transectos de 20 m de largo por 1 m de ancho, para un área total de 320 m². En El Casino se realizaron 3 muestreos en época de lluvia y 3 en la seca, y en cada uno se hicieron 10 transectos de 20 m de largo por 1 m de ancho, para un área total de 200 m², como modificación de la metodología propuesta por Curtis et al. (2004).
A todos los individuos recolectados se les determinó in situ: la especie según Lourie et al. (1999), el sexo gracias al dimorfismo sexual que exhiben los caballitos, la gravidez o no del macho, el estado ovogénico de las hembras por abultamiento abdominal y el Largo Total (LT) o Altura Total (AT), desde el surco medio de la corona hasta el extremo de la cola, con una regla graduada en centímetros (error 0.1 cm). Además se anotó el tamaño del grupo.
Fueron consideradas como hembras todos aquellos individuos con Largo Total (LT) igual o mayor que el del macho más pequeño identificado que fue de 6.4 cm, por lo que todos los individuos menores de esta talla se consideraron indiferenciados.
Se determinaron algunos parámetros fisicoquímicos del agua como: Temperatura, Salinidad. La temperatura se midió con un termómetro (in situ), mientras que la salinidad se determinó con un refractómetro de 1 unidad estándar de salinidad en el laboratorio.
Para verificar la distribución normal de los datos se usó la prueba Kolmogorov-Smirnov con el programa Statistica 6.0 y para comprobar la homogeneidad de varianza de estos se empleó la prueba de Levene. Como los datos no cumplieron con estos requisitos se emplearon análisis estadísticos no paramétricos.
La densidad se analizó para cada especie teniendo en cuenta la estación y época del año. La talla y el estado reproductivo se analizaron por sexo para cada especie. Para la comparación de medias se aplicaron las pruebas de Kruskal Wallis y Man-Whitney. Se aplicó la prueba Chi Cuadrado (Zar, 1996) para determinar las diferencias entre la proporción sexual esperada 1:1 (Lourie et al. 1999). Las diferencias estadísticas fueron consideradas con un nivel de significación del 95% (α = 0.05).
RESULTADOS
Se encontró un total de 37 individuos, de ellos 34 fueron de la especie H. reidi, para un 92% del total en las dos estaciones muestreadas. Solamente aparecieron tres individuos de la especie H. erectus en la zona de Las Cuarenta (Bahía de Cabañas).
H. reidi
Del total de individuos de H. reidi, 12 fueron hembras (35%), 14 machos (41%) y 8 indiferenciados (23%). Esta especie apareció en las dos estaciones y en los macrohábitats de manglar sumergido, estructuras de concretos artificiales y fondo de piedras y algas. Se encontró en un rango de temperatura entre 19-31ºC y en un rango de salinidad entre 30-36 ups.
La densidad media total de la especie fue de 0.0081±0.0017 ind/m2. Se encontraron diferencias significativas entre las estaciones (U = 4613, P = 0.02). El mayor valor de densidad se localizó en la estación de Las Cuarenta (Cuadro 1).

La densidad media total de la especie en época de lluvia fue de 0.0070±0.002 ind/m2, similar a la época seca 0.0095±0.002 ind/m2 (U = 5093, P = 0.53), sin embargo, entre estaciones sí se encontraron diferencias con la época seca, debido a que no aparecieron individuos en El Casino (Fig. 2).
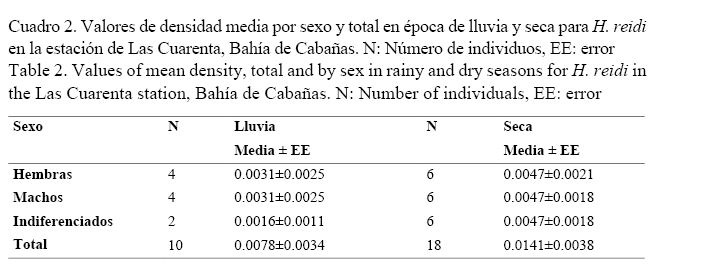
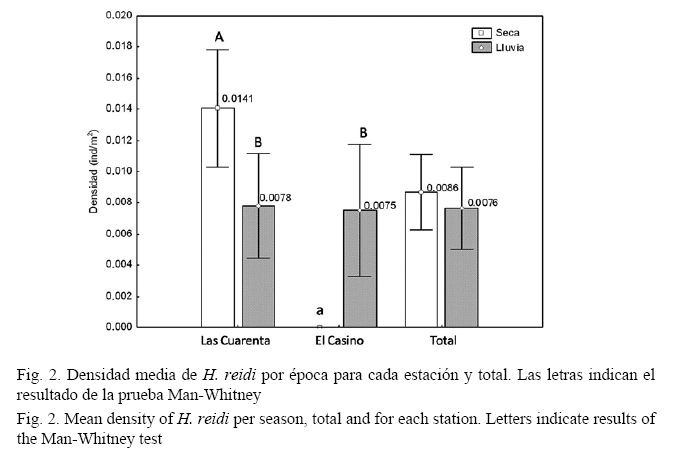
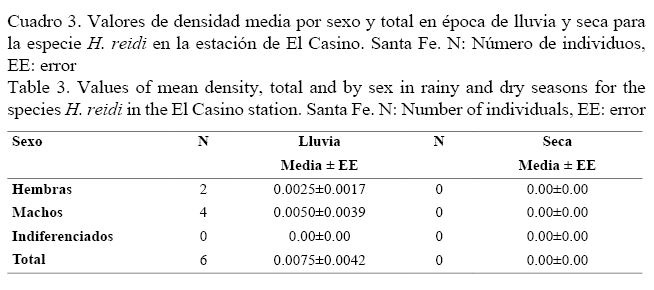
Los valores de densidades medias de H. reidi por sexo y época dentro de cada estación se muestran en los cuadros 2 y 3. En Las Cuarenta, la densidad media en la época seca fue similar a la de lluvia (U = 1831, P = 0.30), mientras que en El Casino solo aparecieron individuos en época de lluvia, como se observa en el cuadro 3.
Se encontró un 28% de machos grávidos que solo aparecieron en los meses de junio y diciembre del 2004 y abril del 2005, por lo que no se pudo definir la época de reproducción. Estos mostraron una talla media de 11.2±0.7 cm (rango de 9.30-12.4 cm).
La proporción sexual fue de 1.16:1 (X2 = 0.15 P > 0.05), similar a la esperada (1:1). En relación con la conducta social, todos los individuos estaban solos y la mayoría se encontraron asociados a diferentes especies de macroalgas, esponjas, ascidias y raíces de mangle rojo.
El largo total promedio de todos los individuos fue de 8.9±0.5 cm (rango de 3.5-14.2 cm). No se encontraron diferencias significativas entre las dos estaciones en cuanto a la talla (U = 41.00, P = 0.05). El menor individuo diferenciado sexualmente fue un macho de 6.4 cm de largo total y la talla del menor macho grávido fue de 9.3 cm de largo total. No se encontraron diferencias entre machos y hembras con respecto al LT (U = 83, P = 0.95).
Aunque no se hizo un estudio detallado sobre los diferentes sustratos utilizados por la especie H. reidi, se puede decir que el mayor número de individuos se encontró en diferentes especies de macroalgas como: C. sertularioides, Caulerpa sp., Penicillum sp. y en las raíces de mangle rojo R. mangle, así como sobre diferentes especies de esponjas.
H. erectus
De los tres individuos encontrados, dos fueron indiferenciados y una era hembra. La especie solo apareció en el biotopo de manglar sumergido de Las Cuarenta. Se localizó en un rango de temperatura entre 19-30ºC y en un rango de salinidad entre 14-40 ups.
La densidad media total de la especie fue de 0.0012±0.0007 ind/m2. La densidad en época de lluvia fue de 0.0008±0.0008 ind/m2 y en época seca de 0.0016±0.0011 ind/m2.
Debido a que no aparecieron machos, no se pudieron determinar la proporción sexual ni la época de reproducción. Todos los individuos se encontraron solos y enganchados en las raíces de mangle rojo. El largo total promedio fue de 6.0±1.0 cm (rango de 4.5-8.0 cm).
DISCUSIÓN
La especie H. reidi se encontró en un rango de temperatura y salinidad mayor que el reportado por Freret-Meurer & Andreata (2008) y a una salinidad menor que la reportada por Mai & Rosa (2009).
La densidad media encontrada en general para esta especie fue menor (0.0081±0.0017 ind/m2) que la reportada por Dias & Rosa (2003); Rosa et al. (2007); Freret-Meurer & Andreata (2008); Mai & Rosa (2009) en diferentes zonas de Brasil, y mayor a la encontrada por Arcos-Pulido & Gómez-Prieto (2004) en Santa Marta, Colombia (Cuadro 4).
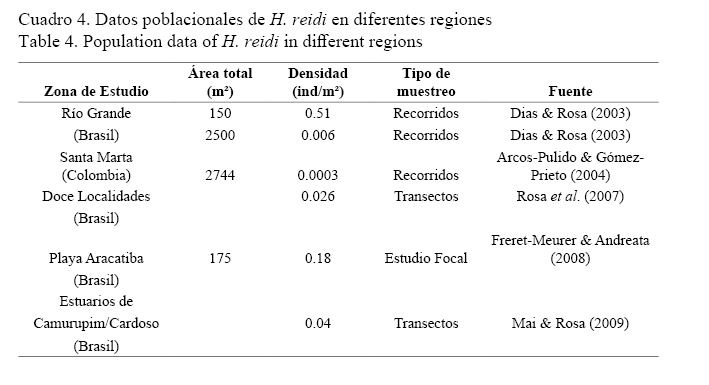
Los valores de densidad media total y por estaciones para H. reidi fueron bajos como se observa en el cuadro 1. Esto coincide con la tendencia general descrita para las especies de caballitos de mar por Lourie et al. (1999; 2004) y Foster & Vincent (2004).
Dias & Rosa (2003) encontraron una marcada diferencia entre las densidades de H. reidi en estructuras artificiales (0.51 ind/m2) y una zona estuarina con manglar, en Río Grande al noreste de Brasil, atribuyendo esta a la atracción que ejercen varios de los microhábitats presentes en el muelle como: macroalgas, esponjas, cnidarios y tunicados. Esto no coincide con lo encontrado en este estudio, donde la densidad en manglar sumergido de Las Cuarenta fue mayor que en las estructuras artificiales de El Casino (0.0037±0.0021 ind/m2).
Es importante destacar que la zona de Las Cuarenta, es un área más protegida y con menos accesibilidad que la de El Casino, que es mucho más urbanizada y con mayor impacto humano, lo que la convierte en una zona más vulnerable a la fácil extracción de caballitos del medio natural (R. Díaz, 2004, com. pers., 8 de agosto). Es posible que esto influya en la abundancia de estas especies.
La densidad encontrada en Las Cuarenta, en biotopo de manglar sumergido, fue mayor que la reportada por Rosa et al. (2007) en las zonas de manglar de Ubatuba y Mamanguape (0.005 y 0.008 ind/m²) y menor que la obtenida en las zonas de Casqueira, Tubarao y Pacoti (0.06 ind/m², 0.055 ind/m² y 0.021 ind/m²) en Brasil.
Los individuos de H. reidi, tanto hembras como machos, presentaron tallas medias menores que las encontradas por Rosa et al. (2007), aunque se debe tener en cuenta que ellos abarcaron un mayor número de localidades y analizaron una mayor cantidad de individuos por sexo.
De forma general, los machos y las hembras de H. reidi presentaron tallas similares. Este resultado no coincide con lo reportado por Rosa et al. (2007) para la misma especie y por Bell et al. (2003) para la especie Hippocampus capensis, donde los machos resultaron mayores que las hembras.
La proporción sexual para H. reidi se corresponde con la frecuencia esperada de 1:1. Esto coincide con Freret-Meurer & Andreata (2008); Mai & Rosa (2009). Según Lourie et al. (1999), esta proporción indica que la estructura social podría ser en parejas, para un comportamiento monógamo de la especie. Dicha organización social es común en muchas especies del género Hippocampus como Hippocampus comes (Perante et al. 2002) e Hippocampus abdominalis (Rosa et al. 2007).
La cantidad de individuos grávidos encontrados en este estudio fue menor que la reportada por Silveira & Fontoura (2004); Rosa et al. (2007). Como solamente se localizaron individuos grávidos en tres meses del año, no se puede determinar la época de reproducción de H. reidi, a diferencia de Rosa et al. (2007) y Freret-Meurer & Andreata (2008), que sí encontraron individuos grávidos en todos los meses, como indicador de que la especie se reproduce todo el año.
Silveira & Fontoura (2004) reportaron picos reproductivos de esta especie en los meses de junio y octubre, mientras que Rosa et al. (2007) los encontraron en los meses de febrero y octubre (meses de verano, hemisferio sur).
La talla de diferenciación sexual, así como el menor macho maduro encontrado en este trabajo, fueron menores que los reportados por Silveira & Fontoura (2004); Rosa et al. (2007) y Mai & Rosa (2009).
El porcentaje de individuos solitarios fue mayor que el reportado por Rosa et al. (2007) y Mai & Rosa (2009). Esto se debe a que dichos autores encontraron varias parejas y grupos de 3 a 7 individuos, mientras que en este estudio todos los individuos estaban solos. Según Claro et al. (2001) es más común localizar a H. reidi solos que formando grupos.
Esta especie se encontró en sustratos similares a los reportados por Dias & Rosa (2003); Rosa et al. (2007) y Mai & Rosa (2009), donde la macroalga (C. sertularioides), el mangle rojo (R. mangle) y el mangle blanco (Laguncularia racemosa), resultaron los más utilizados.
La especie H. erectus se encontró solamente en las raíces de mangle rojo de la estación de Las Cuarenta, en un rango de temperatura menor que el reportado por Teixeira & Musick (2001) y en una salinidad mayor que la reportada por estos autores, corroborando el criterio de que esta especie puede tolerar amplios rangos de salinidad.
Según Teixeira & Musick (2001), H. erectus es más frecuente en aguas profundas donde la vegetación es abundante. Quizás por esa razón se encontraron tan pocos individuos en las estaciones estudiadas, ya que la profundidad de estas no sobrepasa los 2 m, por lo que se sugiere extender este tipo de estudio poblacional en el medio natural a otras áreas de Cuba y realizar muestreos en otros biotopos y a mayor profundidad.
A pesar de que la especie H. erectus se considera similar a H. reidi en cuanto a la historia de vida (Lourie et al. 2004), se encontraron diferencias en su abundancia y hábitat. Esto coincide con Curtis & Vincent (2005), que plantean que especies similares pueden diferir marcadamente en el uso del hábitat y las causas pueden ser: diferencias en la morfología (talla, coloración, presencia de filamentos), conducta y estrategias alimentarias.
El largo total de los individuos encontrados fue menor que el reportado por Baum et al. (2003) y Teixeira & Musick (2001), aunque es importante destacar que estos autores analizaron un número de muestras mucho mayor que el de este estudio.
No se pudieron determinar la proporción sexual ni la época reproductiva de la especie por la ausencia de machos, a diferencia de Teixeira & Musick (2001), que reportaron una proporción sexual diferente a la esperada, y un período reproductivo de mayo a octubre.
CONCLUSIONES
La densidad poblacional de las especies H. reidi e H. erectus, en general y por estaciones, fue baja, aunque la especie H. reidi fue más abundante. La mayor densidad de caballitos se encontró en la estación de Las Cuarenta. No se encontraron diferencias entre la densidad de individuos hallada durante la época de lluvia y seca para ninguna de las dos especies. Los machos y las hembras de H. reidi mostraron tallas similares. Se supone un comportamiento monógamo de la especie H. reidi, debido a la proporción sexual encontrada, muy similar a la proporción de 1:1 esperada.
AGRADECIMIENTOS
Agradecemos a todos los compañeros del grupo de acuariología del Acuario Nacional de Cuba, por su valiosa ayuda en los diferentes muestreos que se realizaron para este trabajo.
BIBLIOGRAFÍA
Arcos-Pulido, M. & Gómez-Prieto, C. (2004). Valoración de la población de Hippocampus reidi en la región de Santa Marta. Recuperado en diciembre 10, 2004, disponible en http://www.sur.iucn.org/listaroja/documentos/Hippocampus.
Baum, J. K., Meeuwig, J. J. & Vincent, A. C. J. (2003). Bycatch of lined seahorses (Hippocampus erectus) in a Gulf of Mexico shrimp trawl fishery. Fish. Bull., 101(4), 721-731.
Bell, E. M., Lockyear, J. F., McPherson, J. M., Marsden, A. D. & Vincent, A. C. J. (2003). First field studies of an Endangered South African seahorse, Hippocampus capensis. Env. Biol. Fish., 67, 35-46.
CITES. (2004). Convención sobre el Comercio Internacional de Especies Amenazadas de Fauna y Flora Silvestres. Recuperado en marzo 18, 2004, disponible en www.cites.org/common/com/AC/20
Claro, R., Lindeman, K. C. & Parenti, L. R. (2001). The Marine Ichthyofauna of Cuba. En R. Claro, K. C. Lindeman & L. R. Parenty (Eds.), Ecology of The Marine Fishes of Cuba (pp. 21-32). Washington, D.C., EE.UU.: Smithsonian Institution.
Curtis, J., Moreau, M. A., Marsden, D., Bell, E., Martin-Smith, K., Samoilys, M. & Vincent, A. (2004). Underwater visual census for seahorse population assessments. Vancouver, B.C., Canada: Project Seahorse, Fisheries Centre.
Curtis, J. M. R. & Vincent, A. C. J. (2005). Distribution of sympatric species along a gradient of habitat complexity in a seagrass-dominated community. Mar. Ecol. Prog. Ser., 291, 81-91.
Dias, T. L. P. & Rosa, I. M. L. (2003). Habitat preferences of a seahorse species, Hippocampus reidi (Teleostei: Syngnathidae) in Brazil. J. Ichthyology Aquat. Biol., 6, 165-176.
Foster, S. J. & Vincent, A. C. J. (2004). Life history and ecology of seahorses: implications for conservation and management. J. Fish. Biol., 65, 1-61.
Freret-Meurer, N. V. & Andreata, J. V. (2008). Field Studies of a Brazilian Seahorse Population, Hippocampus reidi Ginsburg, 1933. Braz. Arch. Biol. Techn., 51(4), 743-751.
Lourie, S. A., Vincent, A. C. J. & Hall, H. J. (1999). Seahorses: an identification guide to the world’s species and their conservation. London, UK: Project Seahorse.
Lourie, S. A., Foster, S. J., Cooper, E. W. T. & Vincent, A. C. J. (2004). A Guide to the Identification of Seahorses. Washington, D.C., EE.UU.: University of British Columbia and World Wildlife Fund.
Lourie, S. A. & Kuiter, R. H. (2008). Three new pygmy seahorse species from Indonesia (Teleostei: Syngnathidae: Hippocampus). Zootaxa, 1963, 54-68.
Mai, A. C. G. & Rosa, I. M. L. (2009). Aspectos ecológicos do cavalo-marinho Hippocampus reidi no estuário Camurupim/Cardoso, Piauí, Brasil, fornecendo subsídios para a criação de uma Área de Proteção Integral. Biota Neotrop., 9(3), 85-91.
Perante, N. C., Pajaro, M. G., Meewig, J. J. & Vincent, A. C. J. (2002). Biology of a seahorse species Hippocampus comes in the central Philippines. J. Fish. Biol., 60, 821-837.
Project Seahorse. (2003). Hippocampus reidi. En IUCN 2003. Red List of Threatened Species. Recuperado en octubre 23, 2003, disponible en http:// www.redlist.org.
Rosa, I. L., Barros, A. T., Xavier, J. H. A. & Cortez, C. S. (2005). Dados populacionais do cavalo-marinho Hippocampus reid Ginsburg, 1933 (Teleostei: Syngnathidae) capturado para fins de aquarismo no nordeste do Brasil. Bol. Técnico-Científico do Cenepe, 13(1), 25-38.
Rosa, I. L., Oliveira, T. P. R., Castro, A. L. C., Moraes, L. E., Xavier, J. H. A., Nottingham, M. C., Dias, T. L. P., Bruto-Costa, L. V., Araújo, M. E., Birolo, A. B., Mai, A. C. G. & Monteiro-Neto, C. (2007). Population characteristics, space use and habitat associations of the seahorse Hippocampus reidi (Teleostei: Syngnathidae), Sociedade Brasileira de Ictiologia. Neotro. Ichthyology, 5(3), 405-414.
Silveira, R. B. & Fontoura, F. N (2004). Reproductive period and size of the first gonad maturation of the Hippocampus reidi sea-horse in the Brazilian Northeast región. Ponencia presentada en el VI International Congress on the Biology of Fishes, Manaus, Brazil.
Teixeira, R. L. & Musick, J. A. (2001). Reproduction and food habits of the lined seahorse, Hippocampus erectus (Teleostei: Syngnathidae) of Chesapeake Bay, Virginia. J. Brazilian Biol., 61(1), 79-90.
UICN. (2004). Red List of Threatened Species. Recuperado en octubre 18, 2004, disponible en www.redlist.org
UICN. (2006). Red List of Threatened Species. Recuperado en mayo 12, 2006, disponible en www.iucnredlist.org
UICN. (2009). Red List of Threatened Species. Recuperado en noviembre 20, 2009, disponible en http://www.iucnredlist.org/apps/redlist.html
UNEP-WCMC CITES. (2006). CITES trade statistics derived from the CITES TradeDatabase, UNEP World Conservation Monitoring Centre, Cambridge, UK. Recuperado en marzo 15, 2006, disponible en http://sea-bov.unep-wcmc.org/citestrade. html
Vincent, A. C. J. (1996). The International Trade in Seahorses. Cambridge, London: TRAFFIC International.
Zar, J. H. (1996). Biostatistical Analysis. New Jersey, EE.UU.: Prentice Hall Inc.

Revista Ciencias Marinas y Costeras está bajo una Licencia Creative Commons Atribución-NoComercial-CompartirIgual 4.0 Internacional.