Rev. Mar. Cost. ISSN 1659-455X. Vol. 4: 105-115, Diciembre 2012.
https://doi.org/10.15359/revmar.4.7
Exploitation and evaluation of the tilapia fishery (Oreochromis spp.) in the Fernando Hiriart Balderrama “Zimapám” Reservoir, Hidalgo, Mexico
Explotación y evaluación de la pesquería de tilapia (Oreochromis spp.) en la represa Fernando Hiriart
Balderrama “Zimapám”, Hidalgo, México
Mario A. Gómez-Ponce1*, María Isabel Gallardo-Berumen2 & Pedro Cervantes-Hernández3
1 Unidad de Servicio el Carmen, Instituto de Ciencias del Mar y Limnología. Universidad Nacional Autónoma de México. Km. 9.5 Carretera Carmen Puerto Real. CP. 24157, Ciudad del Carmen, Campeche, México.
2, 3 Instituto de Recursos, Universidad del Mar, Puerto Ángel, México, Apdo. 47-70902. mgomez@cmarl.unam.mx*
Recibido 18-XI-2011
Aceptado 7-VIII-2012
ABSTRACT
Oreochromis spp. fishing in the Fernando Hiriart Balderrama Reservior began in 1997 and its effects had never been studied before. Schaefer and Fox’s biomass dynamic model were used in order to obtain the first evaluation of this resource and to analyze the presence of record errors (or observation errors) in the annual total catch data set. The number of fishermen was used as the fishing effort unit. Schaefer´s model proved to be statically reliable: Oreochromis spp. was estimated at the maximum population size value of k = 9 000 tons and reproduction rate value at annual r = 0.730. Considering those ecological and biological strategies, Schaefer´s model showed that great biomass levels (predicted biomass) have been produced in the Zimapán reservior; however, only a small part of this biomass was exploited during the analyzed period. The results indicate that tilapia fishery in the Zimapán Reservoir was at equilibrium or was not over-fished between 1997 and 2006 because historical tilapia records observed in Schaefer´s model were far from the k parameter and below the MSY value. With this exploitation type, the fishing effort can increase to up to 7 203 fishermen (level of fishing effort at which the maximum sustainable yield is achieved (fMSY)). However, in order to avoid an increase of the non-controlled fishing effort a similar analysis is recommended with recent data.
Keywords: Dynamic models, evaluation, biomass, tilapia, exploitation.
RESUMEN
La pesca de la tilapia Oreochromis spp. en la presa hidroeléctrica Fernando Hiriart Balderrama comenzó en 1997, esta pesquería nunca ha sido evaluada. Fueron utilizados los modelos dinámicos de biomasa de Schaefer y Fox para obtener la primera evaluación de este recurso y para analizar la presencia de errores de registro (errores de observación) en la captura anual total. El número de pescadores fue usado como la unidad de esfuerzo de pesca. El modelo de Schaefer resultó estadísticamente confiable y para Oreochromis spp. fueron estimados el tamaño máximo de población en k = 9 000 tons y la tasa de reproducción en r = 0.730 anual. Considerando estas estrategias ecológicas y biológicas, el modelo de Schaefer sugirió que grandes niveles de biomasa (biomasa predicha) fueron producidas en la presa Zimapán, pero solo una pequeña parte de esta biomasa fue explotada durante el periodo analizado. Los resultados indicaron que la pesquería de la tilapia en la presa Zimapán estuvo en equilibrio o no fue sobreexplotada entre 1997 y el 2006, porque los registros históricos en el modelo de Schaefer fueron observados muy lejos del parámetro k y por debajo del valor MSY. Con este tipo de explotación, el esfuerzo de pesca podría incrementarse hasta alcanzar 7 203 número de pescadores (nivel de esfuerzo de pesca en el cual se activa el rendimiento máximo sostenible (fMSY)), pero para evitar un incremento no controlado del esfuerzo de pesca, se recomienda hacer un análisis similar integrando datos recientes.
Palabras claves: Modelos dinámicos, evaluación, biomasa, tilapia, explotación.
INTRODUCTION
Water reservoirs have been used to provide and store water, control flooding, generate electricity, produce food, provide jobs and sport fishing (López-Hernández et al. 2007). The Mexican government built the Fernando Hiriart Balderrama Hydroelectric Reservoir (Zimapán Reservoir) in 1995 to produce an average of 1 292.40 GW/year in the state of Hidalgo (Fig. 1). The Zimapán Reservoir is located in the Infiernillo Canyon between the states of Hidalgo and Queretaro and its reservior has a total area of 22.9 km2. The main water sources are the Tula, Moctezuma, Alfajayucan, San Francisco and San Juan rivers (López-Hernández et al. 2007).
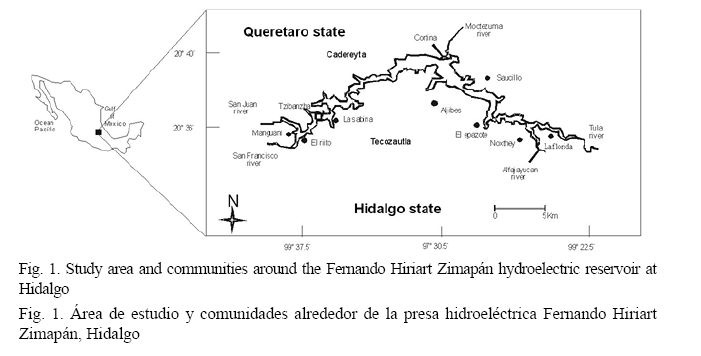
Although tilapia fishing in the Zimapán Reservoir began as a small scale activity in the states of Hidalgo and Queretaro, it was formalized in 1997, increasing tilapia production. Fishing has recently been considered a very important economic activity to provide food and jobs (Hernández-Montaño & Orbe, 2002; López-Hernández et al. 2007). CONAPESCA (2009) reported a 20.45% increase in tilapia production in the states of Hidalgo and Queretaro compared to the one reported in other landlocked Mexican states. Oreochromis spp. is caught in the Zimapán Reservoir with small four meter long fiberglass boats with 15 to 25 horsepower outboard engines. Fishermen use gillnets made of nylon monofilament with a diameter of 0.3 mm or less, minimum mesh size of 11.43 cm, maximum length of 60 m, fall or maximum height of 5 m, and an overlapping between 52% and 65%.
To the best of our knowledge, no other studies about tilapia exploitation have been conducted in the states of Hidalgo and Queretaro. The only reports available discuss water quality in the Zimapán Reservoir (Hernández-Montaño & Orbe, 2002; López-Hernández et al. 2007; Gómez-Ponce et al. 2011). In the present study, tilapia fishing is evaluated from 1997 to 2006, being the first research conducted in the reservoir. Results obtained may be used as a historical point of reference to describe recent biomass changes of Oreochromis spp.
The Schaefer (1954) and Fox (1970) biomass dynamic models (with the observation error) were used in order to evaluate the exploitation (over-fished or non-over-fished) and estimate annual biomass changes. According to Punt & Hilborn (1996), both objectives were developed based on the description of the following parameters: maximum sustainable yield (MSY), level of fishing effort at which the maximum sustainable yield is achieved (fMSY), maximum population size (k), intrinsic growth rate (r), and the catchability coefficient (q).
MATERIALS AND METHODS
The Zimapán Reservoir (20° 35’ - 20° 40’ N and 99° 22’ - 99° 37’ W) has an average surface area of 2 600 ha of water surface, is located at an altitude of 1 870 m above sea level and is fed primarily by the San Juan and Tula rivers. The reservoir is bordered by the municipalities of Zimapán, Tasquillo, and Tecozautla on the western part the State of Hidalgo and by the municipality of Cadereyta Montes within the State of Querétaro. The reservoir has a load capacity of 1.5 million m3 and is considered by the National Fisheries Institute to have great potential for developing tilapia fishing due to its physicochemical conditions and natural and artificial availability of food (López-Hernández et al. 2007).
Total annual catch data sets for Oreochromis spp. were used to estimate catch per unit of effort (CPUE, tons per number of fishermen). This information was obtained during 10 fishing seasons in the Zimapán Reservoir and recorded in the states of Hidalgo and Querétaro between 1997 and 2006. Data sets were provided to the Institute of Maritime and Limnological Sciences (Instituto de Ciencias del Mar y Limnología) from the National Autonomous University of Mexico (UNAM) by the Secretariat of Agriculture, Livestock, Rural Development, Fisheries, and Food from the state of Hidalgo through the records from fishing cooperatives working in the vicinity of the reservoir (SAGARPA).
According to Punt & Hilborn (1996), the Schaefer and Fox models are Bt+1 = [Bt+r·Bt·(1-(Bt/k)-Ct]·Vt and Bt+1 = [Bt+r·Bt·(1-(ln Bt/ln k)-Ct.]·Vt, respectively, where Bt+1 = predicted annual biomass; Bt = present biomass; Ct = present catch; Vt = observation error; k and r are defined as before). Vt = [ln It-ln Ut] (It = present catch per unit of effort; Ut = predicted catch per unit of effort). Bt+1 is “virtial” biomass value (or predicted annual biomass changes) that was available in the Zimapán Reservoir in the year t+1, and on this “virtual” biomass, each year fishermen obtained a real catch value (Ct), which them recorded.
According to Malcolm (2001), the k and r parameters were estimated using the minimum likelihood of logarithmic normal distribution (-ln (k, r/It)), this is:

where SDV is the standard deviation of Vt and n is the total number of fishing seasons analyzed. The catchability coefficient was estimated with:
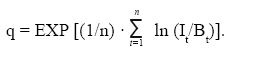
The SDV values obtained were used to evaluate the statistic fit between the It and Ut in both Schaefer´s and Fox´s models. According to Ludwing & Walters (1989), SDV takes values between zero and one. A SDV value near zero guarantees a good statistical fit between It and Ut values; this means that the observation error was not present in all the analyzed fishing seasons or was present only in some of them. Therefore, predicted annual biomass changes of Oreochromis spp. in the Zimapán Reservoir are statistically reliable. A SDV value near one suggests a bad statistic fit between It and Ut values; this means that the observation error was present in all or almost all the analyzed fishing seasons. Therefore, predicted annual biomass changes of Oreochromis spp. in the Zimapán Reservoir are not statistically reliable.
The maximum sustainable yield value and the level of fishing effort at which the maximum sustainable yield is achieved were estimated with: MSY = r·k/4; f =MSY r/2·q. The relationship between fishing effort and tilapia catches was estimated.
According to Chien-Hsiung (2004), the relationships between equilibrium and non-equilibrium in fishery were considered in order to describe the exploitation and the annual biomass changes of Oreochromis spp. The relationships are:
a) When the biomass changes oscillate below MSY value, the system is not over-fished because the level of fishing effort is relatively low. This relationship allows for an increase in the net production without affecting the equilibrium between birth and death rates (the biomass is at equilibrium). Both rates were considered in the r parameter.
b) When the biomass changes have reached or surpassed the MSY value, fishery is generally over-fished by an increase of the fishing effort. This relationship will break the equilibrium between birth and death rates (the biomass is not at equilibrium).
c) The MSY value cannot be greater or equal than the k value.
RESULTS
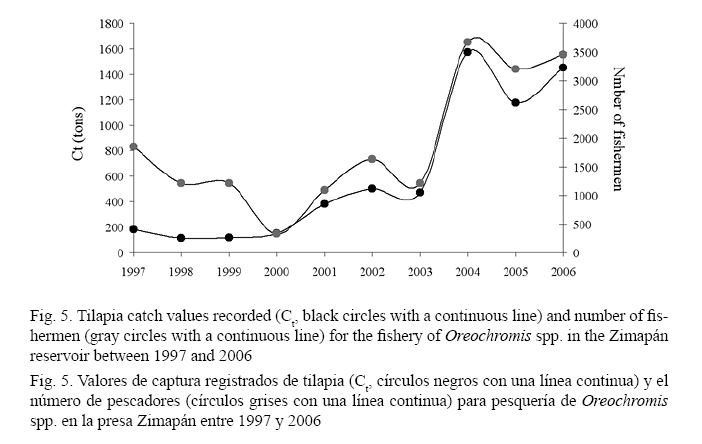
Historical records of tilapia fishing showed that the fishing effort from 1997 to 2000 was equivalent to approximately 200 fishermen while catches decreased from 800 tons captured in 1997 to less than 200 tons. Starting that year, the catch and fishing effort gradually increased to 400 fishermen and 450 tons in 2003, and in 2004 they substantially increased reaching a maximum amount of fishermen at 2 400 and a maximum production of 1 600 tons.
Using the number of fishermen, Schaefer’s model generated a standard deviation value of 0.1788 (Table 1). It and Ut values resulted in a good statistic fit in the 1997, 1998, 2001, 2003, 2004, 2005 and 2006 fishing seasons (70% without recording errors) (Fig. 2). The 1999, 2000 and 2002 fishing seasons showed a bad statistic fit (30% with recording errors) (Fig. 2).


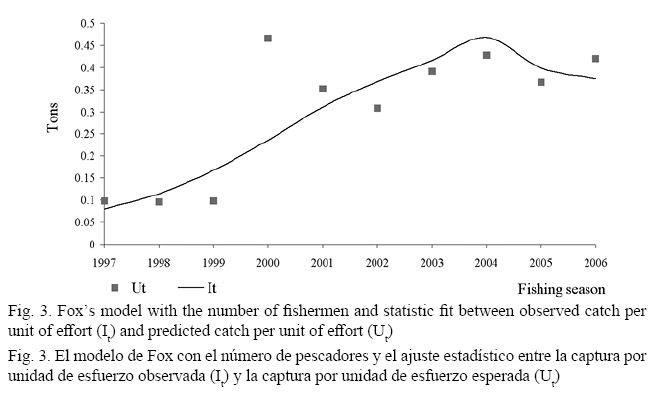
Using the number of fishermen, Fox’s model generated a standard deviation value of 0.3022 (Table 1). The It and Ut values resulted in a good statistic fit in the 1997, 1998, 2001, 2003, 2004, 2005, and 2006 fishing seasons (70% without recording errors) (Fig. 3). The 1999, 2000, 2002 fishing seasons showed a bad statistic fit (30% with recording errors) (Fig. 3). Both objectives were described following Schaefer’s model: Bt+1 = [Bt+0.730·Bt·(1-(Bt/90 000)-Ct]·Vt (Table 1 and Fig. 4).
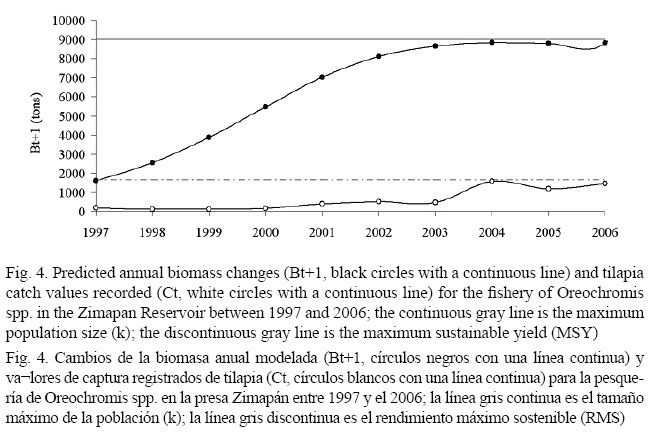
Schaefer’s model showed that predicted annual biomass changes were observed between the MSY and k values (Fig. 4) and that these “virtual” biomass values were never greater than the tilapia catches recorded between 1997 and 2006 (Fig. 4). This result indicates that the Zimapán Reservoir recorded a greater tilapia production, but only a small portion was exploited between 1997 and 2006 (Fig. 4). Following tilapia catch records, two periods of exploitation were observed between 1997 and 2006. In the first period (from 1997 to 2003), tilapia catch was not higher than 600 tons (Fig. 5). In the second period (from 2003 to 2006), tilapia catch increased with a maximum record of 1 573 tons during 2004 (Fig. 5). The end tendency of the fishing effort (number of fishermen) resulted in an equal amount to the one described for tilapia catches (Fig. 5). A directly proportional relationship between the fishing effort and tilapia catches was obtained (r = 0.9310, P = 0.00003).
DISCUSSION
The assessment of fishing resources is critical for managing all fisheries (Penn & Caputi, 1986; Cervantes-Hernández et al. 2006; 2010) since length-cohort and age-cohort data can be carefully analyzed (Jiménez-Badillo, 2004); however, when data is not available, as in the case of a preliminary assessment, the study can be conducted with holistic models, subject to different time scales (Cervantes-Hernández et al. 2006).
The method used to estimate parameters for Schaefer and Fox´s holistic models is the least squares (Sparre & Venema, 1995); however, an increase in the dynamic versions of those parameters (Hilborn & Walters, 1992) has limited their usefulness to manage fisheries, in a way that their application serves only as background (Pérez, 2005; Ibaibarriaga et al. 2008). The least squares method is limited to estimate more than two parameters (Cervantes-Hernández et al. 2006); however, to estimate dynamic parameters, bayesian statistics has been effective in some cases (Morales-Bojórquez et al. 2001; Cervantes-Hernández et al. 2010). For this reason, this investigation has used the minimum plausibility method for obtaining the precise estimation of dynamic parameters r-k-q.
The observation error has in order to evaluate record errors in annual total catch data sets (Punt, 1988) and to generate a reliable statistic fit between the It and Ut (Ludwig & Walters, 1989). The analysis of these errors resulted in an important strategy because the commercial catch-at-age data available for Oreochromis spp. fishery at the Zimapán Reservoir. Based on the results obtained for SDV (Table 1 and Figures 2, 3), Schaefer’s model proved to be statistically reliable and could be used to evaluate exploitation and to estimate annual biomass changes of Oreochromis spp. in the Zimapán Reservoir between 1997 and 2006.
With Schaefer’s model and the observation error, we obtained a reliable statistic fit between the It and Ut (Fig. 2). Standard deviation value of this fit was estimated at 0.1788, suggesting that the predicted annual biomass changes of Oreochromis spp. were statistically reliable than the predicted annual biomass changes obtained with Fox’s model (SDV = 0.3022) (Fig. 3).
We are not aware of other evaluation studies for Oreochromis spp. with this method in Mexico. However, we have gained experience in developing and understanding biomass dynamic models with observation and process errors with other species such as shrimp (Morales-Bojórquez et al. 2001; Cervantes-Hernández et al. 2006). In particular, this study used the experience gained assessing shrimp from the South Pacific to demonstrate how observation errors can affect the evaluation of another type of fishing, in this case, Oreochromis spp. fishing.
Morales-Bojórquez et al. (2001) used the process error in a biomass dynamic model and reported that analyzed commercial catch data had no record errors (or observation errors) but tehy never estimated or demonstrated it. With process error, these authors could not evaluate shrimp fisheries from the Gulf of California, Mexico, because they obtained a “stochastic” behavior in catch per unit of effort records. When catch per unit of effort is “stochastic”, records can not follow a specific trajectory because fishing seasons were not related and an evaluation is not easy to obtain.
According to Hilborn & Walters (1992), the observation error was used in this study to demonstrate that recording errors were present in the annual total catch of Oreochromis spp. and that this error was also considered in the predicted annual biomass changes. A “deterministic” behavior in catch per unit of effort records was observed in Schaefer’s model (Fig. 2). Consequently, the catch per unit of effort records could follow a specific trajectory (statistic fit) where all fishing seasons were related (Fig. 2). With this behavior, fishery was evaluated more easily. Nevertheless, this evaluation showed a bad statistic fit (30% with recording errors) during the 1999, 2000, 2002 fishing seasons (Fig. 2).
In the Zimapán Reservoir, tilapia catch values recorded were never greater than the predicted annual biomass changes of Oreochromis spp. between 1997 and 2006 (Fig. 4). This suggests that the fishing effort never reached an important exploitation level to obtain a point of capture significantly greater than the MSY value obtained. For this reason, we mentioned that this fishery was not over-fished during the studied period. The level of fishing effort at which the maximum sustainable yield is achieved was estimated at 7 203 fishermen (Table 1); however, during all the fishing seasons the total number of fishermen near to the fMSY value was never recorded; the maximum fMSY value was estimated at 1 573 during 2004.
The intrinsic growth rate estimated for Oreochromis spp. resulted at 0.730 annual (Table 1), this value suggested that in the Zimapán Reservoir these species had a fast reproduction rate between 1997 and 2006 (Fig. 4). This result was consistent with fecundity studies reported by Granados-Flores (2006) in the Zimapán Reservoir, this author showed that female tilapia between 17.9 and 25.9 cm of total length had an average production of 2 123 eggs. The reproduction rate of the tilapia life cycle has been described by several authors as high. There have been fertility values ranging between 200 and 3 000 eggs per female depending on the species and the region. Morales (1974) indicated that tilapia females between 31 and 33 cm of total length in the Temascal Miguel Alemán Reservoir, Oaxaca, Mexico, had total oocytes for fish from 1 000 to 1 800. In the Chila lagoon, Basurto (1995) reported that tilapia females between 29 and 34 cm of total length had oocytes for fish production from 888 to 2 945. Jiménez-Badillo (1999) reported that Oreochromis aureus (Steindachner, 1864) females between 13.9 and 28 cm of total length laid or produced an average of 2 123 oocytes in the Adolfo López Mateos Reservoir, Michoacan, Mexico. In the Emiliano Zapata Reservoir, Morelos, Mexico, Peña-Mendoza et al. (2005) observed tilapia females with total length smaller than other females in Mexican Reservoirs (between 15 and 25 cm), which showed a production of oocytes between 243 and 847 (Granados-Flores, 2006). In the case of African tilapias, Anene & Okorie (2008) reported for Tilapia mariae a production of 424 to 2,781 eggs with an average of 1 705 eggs per female, whereas Shalloof-Kariman & Hanan (2008) reported from 289 to 1456 eggs per female for Oreochromis niloticus in a lake in Egypt. Alamilla (2001) reported that Oreochromis spp. is very resistant to live in places with high population density, with severe changes of temperature, salinity and low oxygen concentrations. Bocek (2003) indicated that Tilapia can feed on a wide range of natural and artificial food. Considering these ecological aspects and reproductive strategies, we can understand the high values of the k parameter (9 000 tons) present in the Zimapán Reservoir (Table 1 and Fig. 4). Several authors have reported an ecological relationship between the k and r parameters (Krebs, 1985; Beltrán-Álvarez et al. 2009).
Based on the results from this study, we conclude that tilapia fishing in the Zimapán Reservoir was at equilibrium or was not over-fished during the analyzed period (1997 to 2004) because in all fishing seasons, tilapia catches recorded in Schaefer´s model were observed far from the k parameter and below the MSY value.
ACKNOWLEDGEMENTS
We would like to thank Adriana Vallarino Moncada, María de los Ángeles Cauich García, Esther Diupotex Chong, Hernán Álvarez Guillén, Andrés Reda Deara, Derek Brockett and anonymous reviewers for their valuable time and comments regarding this paper.
REFERENCES
Alamilla, H. (2001). Cultivo de tilapia. City of Mexico, Mexico, D. F.: Techni-Campo.
Anene, A. & Okorie, P. U. (2008). Some aspects of the reproductive biology Tilapia mariae (Boulenger 1901) in a small lake in southeastern Nigeria. Afri. J. Biotech., 7(14), 2478-2482.
Basurto, M. (1995). Algunos aspectos reproductivos de la tilapia Oreochromis niloticus en la Laguna de Chila, Veracruz. Biotam., 6(3) 32-41.
Beltrán-Álvarez, R., Sánchez, P. J., Valdez, G. L. & Salas, A. O. (2009). Edad y crecimiento de la mojarra Oreochromis aureus (Pisces: Cichlidae) en la Presa Sanalona, Sinaloa, México. Rev. Biol. Trop., 58, 325-338.
Bocek, A. (2003). Reproductive biology of Oreochromis niloticus. Alabama, USA.: International Center for Aquaculture and Aquatic Environment.
Cervantes-Hernández, P., Ramos-Cruz, S. & Gracia, A. (2006). Evaluación del estado de la pesquería de camarón en el Golfo de Tehuantepec. Hidrobiol., 16(3), 233-239.
Cervantes-Hernández, P., Flores-Gómez, A., Serrano-Guzmán, S. J., Ramos-Cruz, S. & Gallardo-Berumen, M. I. (2010). Historical exploitation and evaluation of brown shrimp fishery Farfantepeneaeus californiensis (Decapoda, Dendrobranchiata) in the Gulf of Tehuantepec, Oaxaca, Mexico. Pan. Amer. J. Aquatic. Sci., 5(4), 486-494.
Chien-Hsiung, W. (2004, August). Improvement of the Schaefer model and its application. Paper presented in the 17th Meeting of the Standing Committee on Tuna and Billfish, Taipei, Taiwan.
CONAPESCA. (2009). Anuario estadístico de Pesca. City of Mexico, Mexico, D. F.: SAGARPA.
Fox, W. (1970). An exponential yield model for optimizing exploited fish populations. Trans. Amer. Fish. Soc., 99, 80-88.
Gómez-Ponce, M. A., Granados-Flores, K., Padilla, C., López-Hernández, M. & Núñez Nogueira, G. (2011). Edad y crecimiento del híbrido de tilapia Oreochromis niloticus x Oreochromis aureus (Perciformes: Cichlidae) en la represa “Zimapán” Hidalgo, México. Rev. Biol. Trop., 59(2), 761-770.
Granados-Flores, K. (2006). Estudio preliminar de edad y crecimiento de la tilapia (Oreochromis niloticus x Oreochromis aureus), con base en escamas, en la presa Fernando Hiriart Valderrama “Zimapán”. Unpublished thesis, Universidad Nacional Autónoma de México, México.
Hernández-Montaño, D. & Orbe, A. (2002). Evaluación pesquera de la presa Fernando Hiriart Balderrama (Zimapán) Hidalgo-Querétaro, México. Hidalgo, México: INP.
Hilborn, R. & Walters, C. J. (1992). Quantitative fisheries stock assessment. Choice, dynamics and uncertainty. New York, USA: Chapman & Hall.
Ibaibarriaga, L., Fernández, C., Uriarte, A. & Roel, B. A. (2008). A two-stage biomass dynamic model for Bay of Biscay anchovy: a Bayesian approach. ICES J. Mar. Sci, 65, 191-205.
Jiménez-Badillo, L. (1999). Análisis de la pesquería de tilapia Oreochromis spp. (Pisces: Cichlidae) en la Presa Adolfo López Mateos, Michoacán-Guerrero. Unpublished doctoral dissertation. Universidad Nacional Autónoma de México, México.
Jiménez-Badillo, L. (2004). Application of holistic and analytical models for the management of tilapia fisheries in reservoirs. Hidrobiol, 14(1), 61-68.
Krebs, C. J. (1985). Ecología, estudio de la distribución y la abundancia. City of Mexico, Mexico, D. F.: Harla.
López-Hernández, M., Ramos, G., Gómez-Ponce, M. A., Figueroa-Torres, M. & Carranza-Fraser, J. (2007). Aspectos limnológicos, de impacto ambiental y pesqueros en la presa hidroeléctrica Zimapán, Hidalgo-Querétaro. In J. Arredondo, G. Díaz & J. Ponce (Eds.), Limnología de presas mexicanas (pp. 801-820). City of Mexico, Mexico, D. F.: AGT. Editores.
Ludwig, D. & Walters, C. J. (1989). A robust method for parameter estimation from catch and effort data. Can. J. Fish. Aquat. Sci., 46, 137-144.
Malcolm, H. (2001). Modelling and quantitative methods in fisheries. London, England: Chapman & Hall.
Morales, A. (1974). El cultivo de la tilapia en México. Datos biológicos. City of Mexico, Mexico, D. F.: INP.
Morales-Bojórquez, E., López-Martínez, J. & Hernández-Vázquez, S. (2001). Modelo dinámico de captura y esfuerzo para el camarón café Farfantepenaeus californiensis (Holmes) del Golfo de California, México. Cienc. Mar., 27(1), 105-124.
Penn, J. W. & Caputi, N. (1986). Spawning stock-recruitment relationships and environmental influences on the tiger prawn (Penaeus esculentus) fishery in Exmouth Gulf, Western Australia. J. Mar. Fresh. Res., 37, 491-505.
Peña-Mendoza, B., Gómez-Márquez, J. L., Salgado-Ugarte, I. H. & Ramírez-Noguera, D. (2005). Reproductive biology of Oreochromis niloticus (Perciformes: Cichlidae) at Emiliano Zapata dam, Morelos, Mexico. Rev. Biol. Trop., 53(3-4), 515-522.
Pérez, E. P. (2005). Un modelo simple para describir la dinámica de la biomasa del camarón nailon Heterocarpus reedi en Coquimbo, Chile. Invest. Mar., 33(2), 131-142.
Punt, A. E. (1988). Model selection for dynamics of southern Africa hake resources. Unpublished Master’s thesis, Department of Mathematics, University of Cape Town, South Africa.
Punt, A. E. & Hilborn, R. (1996). Biomass dynamic models. User`s Manual. Rome, Italy: FAO.
Schaefer, M. B. (1954). Some aspects of the dynamics of populations important to the management of the commercial marine fisheries. Inter. Amer. Trop. T., 1(2), 27-56.
Shalloof-Kariman, A. & Hanan, M. M. (2008). Investigation on some aspects of reproductive biology in Oreochromis niloticus (Linnaeus, 1757) Inhabited Abu-zabal Lake, Egypt. Glob. Vet., 2(6), 351-359.
Sparre, P. & Venema, S. C. (1995). Introducción a la evaluación de recursos pesqueros tropicales. Manual. Rome, Italy: FAO.