Rev. Mar. Cost. ISSN 1659-455X. Vol. 6: 115-133, Diciembre 2014.
DOI: http://dx.doi.org/10.15359/revmar.6.8
Efectos de El Niño y La Niña sobre el comportamiento del microfitoplancton marino y las variables fisicoquímicas durante el 2008 a 2010 en el Golfo de Nicoya, Costa Rica
Effects of El Niño y La Niña over marine microphytoplankton behavior and physical-chemical variables during the 2008-2010 period in the Gulf of Nicoya, Costa Rica
Emilia Calvo Vargas1*, Jorge Boza Abarca1 y Karen Berrocal Artavia1
1 Estación de Biología Marina Juan Bertoglia Richards, Escuela de Ciencias Biológicas, Universidad Nacional, Puntarenas, Costa Rica. emilia.calvo.vargas@una.cr* y ecalvo09@gmail.com
Recibido: 30 de marzo de 2014
Corregido: 25 de junio de 2014
Aceptado: 17 de julio de 2014
RESUMEN
El Niño 2009 fue responsable de una estación lluviosa de baja intensidad y de valores promedios anuales de temperatura del agua, sólidos totales, salinidad y oxígeno disuelto más altos. Las densidades medias de diatomeas y dinoflagelados disminuyeron durante el 2009, mientras que las densidades medias de cianobacterias y silicoflagelados permanecieron similares en La Niña 2008 y La Niña 2010. En el Golfo de Nicoya, este evento causó cambios significativos en la abundancia y/o la presencia-ausencia de las diferentes especies que forman la comunidad microfitoplanctónica de acuerdo con: (1) disminución de la densidad de los dinoflagelados: G. catenatum, A. fraterculus, A. monilatum, D. caudata, N. scintillans, C. furca, Tipo 1, C. fusus, P. conicum, G. verior, C. tripos, L. polyedrum, C. lineatum, G. polygramma, y la diatomea S. costatum; (2) aumento de la densidad de los dinoflagelados: C. macroceros, P. kofoidii, P. micans, C. furca, y las cianobacterias: T. erythraeum y M. elegans; (3) presencia exclusiva de dinoflagelados: G. instriatum, Ceratocorys sp., P. bipes, C. massiliense, G. instriatum, P. balticum, y la diatomea Aulacodiscus sp.; y (4) ausencia de: C. lineatum, C. trichoceros, O. steinii, Alexamdrium sp. y C. macroceros. La presencia exclusiva de determinadas microalgas y la composición de la comunidad microfitoplanctónica son consideradas indicadores biológicos marinos potenciales de los eventos El Niño y La Niña en el Golfo de Nicoya. Recomendamos continuar en el futuro con el estudio de las condiciones hidrográficas y composición del fitoplancton, haciendo énfasis en las especies consideradas como indicadores biológicos y los nutrientes principales.
Palabras claves: El Niño, La Niña, microfitoplancton, bioindicadores, Golfo de Nicoya.
ABSTRACT
The 2009 El Niño phenomenon was responsible for a rainy season of low intensity as well as higher annual average water temperatures, total solids, salinity and dissolved oxygen. Average densities of diatoms and dinoflagellates decreased during 2009, while average densities of cyanobacteria and silicoflagellates were similar in the 2008 and 2010 La Niña phenomena. In the Gulf of Nicoya, this phenomenon caused significant changes in abundance and/or presence-absence of different species comprising the microphytoplankton community based on: (1) Decreased density of dinoflagellates: G. catenatum, A. fraterculus, A. monilatum, D. caudata, N. scintillans, C. furca, Type 1 C. fusus, P. conicum, G. verior, C. tripos, L. polyedrum, C. lineatum, G. polygramma, and diatom S. costatum; (2) Increased density of dinoflagellates: C. macroceros, P. kofoidii, P. micans, C. furca, and cianobacteria T. erythraeum and M. elegans; (3) Exclusive presence of dinoflagellates: G. instriatum, Ceratocorys sp., P. bipes, C. massiliense, G. instriatum, P. balticum, and diatom Aulacodiscus sp.; and (4) Absence of C. lineatum, C. trichoceros, O. steinii, Alexamdrium sp., and C. macroceros. The exclusive presence of some microalgae and the microphytoplankton community composition are considered potential marine biological indicators of El Niño and La Niña in the Gulf of Nicoya. Further studies of the hydrographic conditions and the phytoplankton composition are recommended, with special focus on those species considered as biological indicators and main nutrients.
Keywords: El Niño, La Niña, microphytoplankton, bioindicators, Golfo de Nicoya.
INTRODUCCIÓN
El Niño (EN) es conocido como un fenómeno atmosférico que causa el aumento de la temperatura superficial del mar (TSM) a lo largo del Pacífico ecuatorial (Todd et al. 2011; Gajardo et al. 2013), afectando las costas de Chile, Perú, Ecuador, Colombia, Centroamérica y México (Tarazona et al. 1999; Rodríguez, 2004; Tapia, 2007; Brugnoli-Olivera & Morales-Ramírez, 2008; García et al. 2008; Gaxiola-Castro et al. 2008). Por el contrario, La Niña (LN) provoca la disminución de la TSM en las mismas áreas (Escalante et al. 2013; Gajardo et al. 2013).
Los efectos directos de EN incluyen cambios ambientales como: calentamiento de la capa de mezcla, aumento del nivel marino, aumento en el flujo geostrófico hacia los polos, cambios en los patrones de circulación, vientos más débiles, hundimiento de la termoclina, disminución en el enriquecimiento del agua y disminución en la producción primaria (fitoplancton) (Escalante et al. 2013; Gajardo et al. 2013).
Durante un evento de LN afloran aguas subsuperficiales más frías que las normales en el centro y este del Océano Pacífico ecuatorial (García et al. 2008; Escalante et al. 2013; Gajardo et al. 2013), provocando que la corriente fría de Humbolt fluya a lo largo de la costa sudamericana, transportando aguas ricas en nutrientes que propician un aumento de la producción primaria (Gajardo et al. 2013). Ambos fenómenos atmosféricos alteran los patrones de precipitación, por lo general, durante un evento de EN la precipitación disminuye, mientras que en LN aumenta (Magaña et al. 2003). Debido a las variaciones en las condiciones ambientales que provocan estos fenómenos, varios grupos taxonómicos han sido catalogados como indicadores de EN y LN, siendo los principales las diatomeas, los dinoflagelados (Ochoa, 2003; Palomares-García et al. 2003; Rodríguez, 2004; Rojas & Ortiz, 2007; Tapia, 2007) y foraminíferos planctónicos (Gajardo et al. 2013).
Las costas de Costa Rica se han visto afectadas por ambos fenómenos, durante EN 1997 se registraron variaciones en las concentraciones de nutrientes, en la abundancia de fitoplancton y el incremento de la temperatura en el Golfo de Nicoya, provocando cambios en la estructura de las poblaciones planctónicas en la zona interna del golfo (Brugnoli-Olivera & Morales-Ramírez, 2008).
En el Golfo de Nicoya se mantuvo un monitoreo mensual de las condiciones fisicoquímicas y biológicas (microfitoplancton) durante el 2008 al 2010. Según El Índice de El Niño Oceánico (INO) de la NOAA (2011), durante el 2008 ocurrió un LN con una intensidad moderada, en el 2009 ocurrió un EN con una intensidad fuerte, y en el 2010, nuevamente, fue LN con intensidad moderada (Boening et al. 2012).
Por lo anterior, el objetivo de este estudio fue relacionar el cambio temporal de las variables fisicoquímicas y biológicas (microfitoplancton) durante el período en que se registraron estos tres eventos atmosférico-oceanográficos, con el fin de que sus variaciones en el Golfo de Nicoya sirvan como indicadores de este tipo de fenómenos climáticos.
MATERIALES Y MÉTODOS
Descripción del área de estudio
El Golfo de Nicoya (Fig. 1), situado en la costa pacífica de Costa Rica, es considerado un estuario, cuyos ecosistemas litorales y ríos conforman una base rica de recursos naturales de gran valor económico para el país (Castro & León, 1998). El golfo tiene una longitud de aproximadamente 80 Km, un ancho de 50 Km y un área de 1 550 Km2 (León-Coto et al. 1998). Se divide en la parte interna, desde la desembocadura del río Tempisque hasta Puntarenas, caracterizada por un litoral de manglar y una profundidad no mayor a los 20 m; y la parte externa con influencia de los ríos Grande de Tárcoles y Barranca, donde la costa alterna zonas rocosas con playas arenosas y las profundidades van de los 25 m hasta más de 100 m (Calvo, 2002).
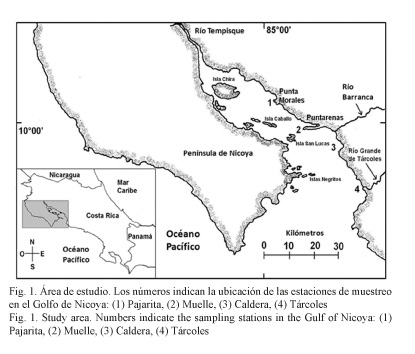
Se establecieron cuatro estaciones de muestreo en el Golfo de Nicoya (Fig. 1): (1) Isla Pajarita (Pajarita) (10° 5’ 30’’ N y 84° 59’ 80’’ O) recibe la influencia del río Tempisque y caracteriza la parte interna del golfo; (2) Muelle de Puntarenas (Muelle) (9° 58’ 25’’ N y 84° 49’ 76’’ O), en la parte media del golfo, donde confluyen las aguas internas y las externas; (3) bahía Caldera (Caldera) (9° 54’ 90’’ N y 84° 43’ 40’’ O), con influencia de la descarga del río Barranca; y (4) Río Grande de Tárcoles (Tárcoles) (9° 44’ 60’’ N y 84° 38’ 70’’ O), con influencia del Río Grande de Tárcoles, ambas estaciones ubicadas en la parte externa del golfo e influenciadas por aguas oceánicas.
Cada quince días se realizó un muestreo en cada una de las estaciones durante el período comprendido entre enero de 2008 a diciembre de 2010. Se midieron in situ a 1 y 5 m de profundidad, la temperatura del agua (°C), la salinidad (UPS), los sólidos totales (g L-1), el oxígeno disuelto (mg L-1) y la profundidad del disco de Secchi (m). Todas las variables se midieron con una sonda multiparamétrica (modelo YSI Incorporated, YSI 556 MPS). Además, en cada estación se tomaron muestras de agua a 1 y 5 m de profundidad utilizando una botella oceanográfica Niskin de 5 L de capacidad. La concentración celular en las muestras de agua se determinó utilizando el método Utermöhl (Reguera, 2002), con columnas de 10 mL. Las muestras se sedimentaron por al menos 12 horas antes de realizar los análisis. En el laboratorio y empleando un microscopio invertido (Nikon Eclipse TE 2000-S) se identificaron las especies de microalgas y el número de individuos por especie y muestra (Cél. L-1, n = 2). Las microalgas fueron agrupadas como diatomeas, dinoflagelados, cianobacterias y silicoflagelados, y se determinó su abundancia relativa en las muestras (Porcentaje mensual). Con los valores promedios anuales de las especies individuales se determinaron aquellas especies que disminuyeron o aumentaron su densidad, o que estuvieron presentes o ausentes durante el período de estudio. Los resultados se complementaron con datos de precipitación (mm) suministrados por el Instituto Meteorológico Nacional de Costa Rica, y de épocas de eventos atmosféricos de EN y LN (NOAA, 2011) para los tres años.
Para determinar las variaciones mensuales de los parámetros fisicoquímicos y del microfitoplancton, se obtuvieron promedios mensuales por estación de muestreo. Los datos fueron procesados y analizados estadísticamente a escala espacio-regional (Fig. 1), global (considerando al golfo como una unidad) y temporal (mensual y anual), y se compararon con las variaciones anuales de los eventos de LN del 2008 y el 2010, y el evento de EN 2009.
Para establecer las diferencias estadísticas en las variables estudiadas entre cada evento (LN 2008, EN 2009 y LN 2010), se realizaron un análisis de varianza, ANDEVA (SPSS, 2006) y una prueba de comparaciones múltiples de Tuckey DHS (Diferencia Honestamente Significativa con P ˂ 0.05).
Resultados
La variación mensual de la TSM (Fig. 2A) mostró valores bajos al inicio del año, incrementos con la entrada de la época lluviosa (de mayo a noviembre) y disminuciones en los últimos meses del año, al igual que a 5 m. Las temperaturas mensuales durante el segundo semestre de 2009 fueron mayores con respecto a las observadas en el 2008 y el 2010. En el 2009, el promedio anual de la TSM fue de 28.73 ± 0.17°C (Cuadro 1), 0.60°C más alto que en el 2008 y 0.49°C superior que en el 2010. A 5 m de profundidad, el promedio anual en el 2009 fue de 28.29 ± 0.18°C, siendo superior que en el 2008 por 0.74°C, y por 0.79°C en el 2010. Estas diferencias fueron estadísticamente significativas (P ˂ 0.05).
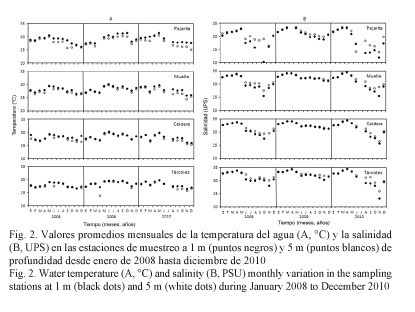
La salinidad (Fig. 2B) fue alta en el primer semestre de todos los años de estudio, disminuyendo con la entrada de la época lluviosa e incrementándose a finales de cada año. Las variaciones en el segundo semestre fueron más fuertes en el 2008 y el 2010, con respecto al 2009. La salinidad a 1 m fue mayor en el 2009 (32.18 ± 0.20 UPS) que en el 2008 y el 2010 (2.37 UPS y 2.09 UPS menores significativamente (P ˂ 0.05), respectivamente). A 5 m de profundidad, el promedio anual en el 2009 alcanzó 32.47 ± 0.16 UPS, siendo inferior en el 2008 (por 1.25 UPS) y en el 2010 (por 1.53 UPS) (P ˂ 0.05, Cuadro 1).
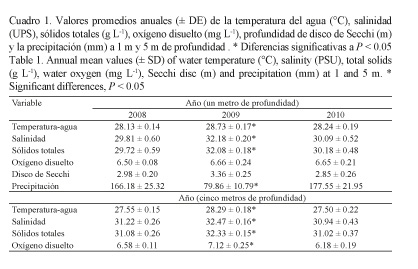
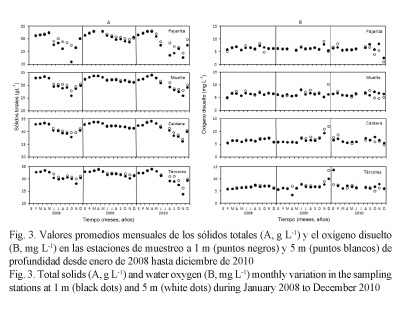
Los sólidos totales (Fig. 3A) tuvieron un comportamiento similar a la salinidad con valores altos al inicio de año (época seca, de diciembre a abril) y bajos a mediados y finales de año, principalmente en el 2008 y el 2010. Los valores en el 2009 fluctuaron levemente en todas las estaciones. Los sólidos totales anuales a 1 m fueron 32.08 ± 0.18 g L-1 (2009), valor superior al del 2008 por 2.36 g L-1 y al del 2010 por 1.90 g L-1 (P ˂ 0.05); y a 5 m fue de 32.33 ± 0.15 g L-1 en el 2009, con diferencias de 1.25 g L-1 y 1.31 g L-1 para el 2008 y el 2010, respectivamente (P ˂ 0.05, Cuadro 1).
El oxígeno disuelto (Fig. 3B) fue similar durante todo el 2008 y el 2010, mientras que en el 2009 se observó un incremento fuerte en todas las estaciones a finales de año. Similar a las variables anteriores, el promedio anual del oxígeno disuelto fue superior durante el 2009, en ambas profundidades.
Los valores promedios mensuales de las cuatro estaciones (Golfo de Nicoya) para cada variable fueron comparados con la profundidad del disco de Secchi, la precipitación y el INO (Fig. 4). Durante el primer semestre de cada año de estudio, la profundidad del disco de Secchi fue alta, disminuyendo con la entrada de la época lluviosa en el segundo semestre e incrementándose al finalizar cada año. El promedio anual de la profundidad del disco de Secchi fue menor en 0.38 m (2008) y en 0.51 m (2010), con respecto al valor del 2009 (3.36 ± 0.25 m) (P ˂ 0.05, Cuadro 1).
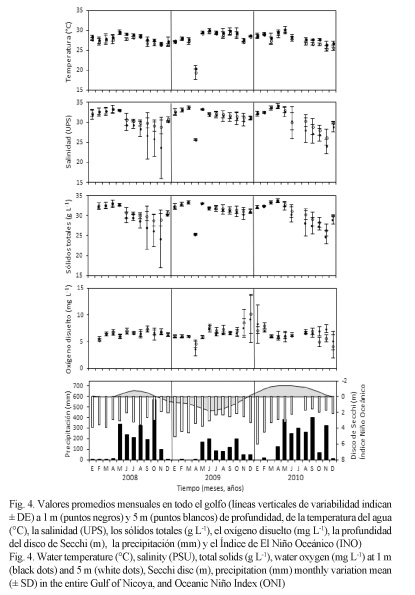
La época lluviosa en el 2008 y el 2010 fue más intensa que en el 2009 (Cuadro 1), y el INO presentó valores negativos durante estos años, mientras que durante el 2009 fueron positivos (Fig. 4). En general, el promedio anual de precipitación fue más alto en el 2008 (166.18 ± 25.32 mm) y el 2010 (177.55 ± 21.95 mm) y menor en el 2009 (79.86 ± 10.79 mm), el cual fue significativamente diferente (P ˂ 0.05).
Durante los tres años de estudio se identificaron taxonómicamente 2 especies de diatomeas, 54 especies de dinoflagelados, 2 especies de cianobacterias y una especie de silicoflagelado.
Las densidades promedios anuales de las diatomeas fueron mayores durante el 2008 y el 2010, disminuyendo en el 2009 (P ˂ 0.05, Cuadro 2); esta variación es más evidente a 1 m de profundidad (Fig. 5A) que a 5 m (Fig. 5B). Los dinoflagelados presentaron una variación similar a la de las diatomeas a 1 m de profundidad (P ˂ 0.05, Cuadros 2 y 3).
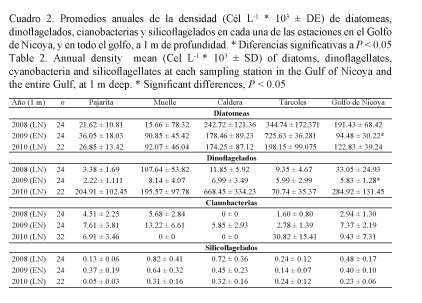

Los grupos de cianobacterias (Cuadros 2 y 3), a 1 y 5 m de profundidad, tendieron a aumentar su densidad en el promedio anual a partir de 2009, y llegaron a valores pico en el 2010; los silicoflagelados, por el contrario, disminuyeron hasta alcanzar valores promedios anuales mínimos en el 2010.
Las especies con densidades bajas en el 2009 con respecto a los otros años, a 1 m de profundidad (Cuadro 4), fueron los dinoflagelados: Gymnodinium catenatum, Alexandrium fraterculus, A. monilatum, Dinophysis caudata, Noctiluca scintillans, Ceratium furca, Tipo 1, C. fusus, Protoperidinium conicum, Gonyaulax verior, C. tripos, Lingulodinium polyedrum, y la diatomea Skeletonema costatum; y a 5 m de profundidad (Cuadro 5): G. catenatum, A. fraterculus, A. monilatum, N. scintillans, Tipo 1, C. lineatum, G. verior, P. conicum, C. trichoceros, L. polyedrum, S. costatum y Gonyaulax polygramma.
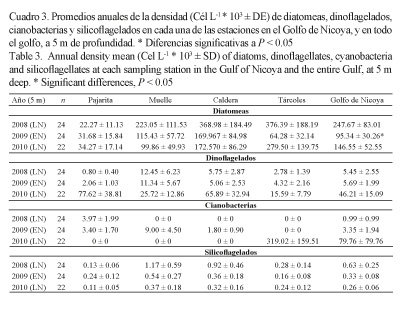
Las especies que aumentaron su densidad en el 2009 a 1 m de profundidad fueron los dinoflagelados C. macroceros, Polykrikos kofoidii, y las cianobacterias Trichodesmium erythraeum y Merismopedia elegans; y a 5 m de profundidad los dinoflagelados Prorocentrum micans y C. furca (Cuadros 4 y 5).
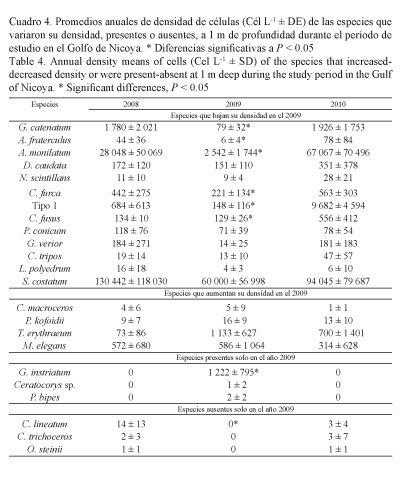
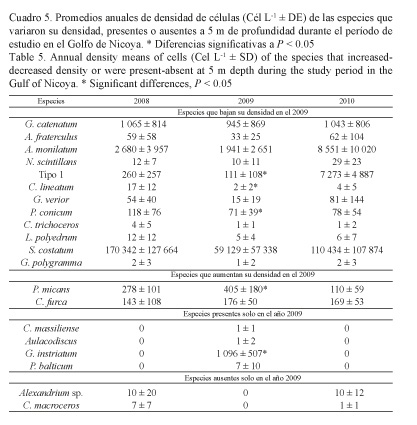
Las especies presentes en el 2009, pero ausentes en el 2008 y el 2010, a 1 m de profundidad, fueron los dinoflagelados: Gyrodinium instriatum, Ceratocorys sp. y Podolampas bipes; mientras que a 5 m de profundidad fueron: C. massiliense, G. instriatum y Prorocentrum balticum, y la diatomea Aulacodiscus sp. (Cuadros 4 y 5).
Por último, se identificaron especies que estuvieron ausentes en el 2009, pero que sí aparecieron en las muestras de 2008 y 2010. A 1 m de profundidad fueron los dinoflagelados: C. lineatum, C. trichoceros y Ornithocercus steinii; y a 5 m de profundidad: Alexandrium sp. y C. macroceros (Cuadros 4 y 5).
Discusión
De acuerdo con el INO de la NOAA (2011), durante el período de estudio ocurrieron tres eventos en el Pacífico ecuatorial frente a las costas de América: LN leve en el 2008, EN fuerte en el 2009 y LN moderada en el 2010 (Fig. 4). Esta sucesión de eventos permitió evaluar su efecto sobre el comportamiento de las variables fisicoquímicas y el microfitoplancton del Golfo de Nicoya.
Durante EN 1997 se determinó en la parte interna del Golfo de Nicoya una temperatura superficial máxima de 31.5°C (abril), y una mínima de 28°C (marzo y octubre) (Brugnoli & Morales, 1999; Morales-Ramírez & Brugnoli-Olivera, 2001; Brugnoli-Olivera & Morales-Ramírez, 2008). Sin embargo, esta es la primera vez que se determina la disminución de la TSM durante eventos LN (2008 y 2010). Durante EN 2009 se registró un aumento de la TSM con respecto a los eventos LN 2008 y 2010, similar a lo observado en otras zonas. En México, durante EN 1982-1983, se observaron diferencias de +2.6°C, y en EN 1997-1998, en la misma área, las diferencias de la TSM superaron los 4.4°C (Badan, 2003; Palomares-García et al. 2003). En la costa suroeste de México (EN 1997-1998),- se determinó un aumento de la TSM de 7.6°C con respecto a la de enero de 1996 (Filonov et al. 2003), y en la costa sur-occidental (período 1996-1999) se midieron diferencias de 3 a 5°C con respecto a un período normal (Aguirre-Gómez et al. 2003; Badan, 2003).
Durante el evento EN 1997-1998, en el Pacífico colombiano, la TSM se incrementó entre 2.2 y 2.6°C con respecto a la normal (27.0°C y 27.4°C); el aumento en la temperatura fue provocado por el flujo de aguas cálidas de la contracorriente norecuatorial (García et al. 2008). Recientemente, Escalante et al. (2013) encontraron en el Golfo de California diferencias significativas en la TSM entre los eventos EN (25.50°C) y LN (24.19°C).
En este estudio, la salinidad en el 2009 presentó un valor promedio anual de 32.18 UPS, superior al de 2008 y 2010 (Cuadro 1), siendo en abril de 2009 el valor máximo de 34.64 UPS a 5 m de profundidad. Durante EN 1997-1998, se midieron en la parte interna del Golfo de Nicoya salinidades máximas de 35 UPS (diciembre) (Brugnoli & Morales, 1999; Brugnoli-Olivera & Morales-Ramírez, 2008). En la costa suroeste de México, durante EN 1998 se midió una salinidad de 34 UPS (enero), 0.5 UPS menor a la de enero de 1996 (evento LN) (Filonov et al. 2003). En Colombia, durante EN 1991-1992, se detectó un incremento de la salinidad de hasta 34 UPS (Castillo & Vizcaíno, 1993), que García et al. (2008) durante EN 1997-1998, registraron valores más bajos (24-30 UPS), entretanto durante LN, la salinidad fue superior (31.6 UPS). En este estudio, probablemente el evento EN 2009 causó el aumento de la salinidad del agua en el Golfo de Nicoya por más de 2 UPS.
En el presente estudio, el oxígeno disuelto aumentó durante EN 2009, siendo más evidente la diferencia a finales del año, mientras que en el 2008 y el 2010 fue menor. Durante EN 1997-1998, en el Golfo de Nicoya se determinó un oxígeno disuelto máximo de 8 mg L-1 (noviembre). Este comportamiento podría estar relacionado con el ingreso al golfo de agua supersaturada subsuperficial, con la finalización de la época lluviosa e inicio de la época seca (Brugnoli-Olivera & Morales-Ramírez, 2008) y con la estratificación provocada por la composición de las aguas costeras que entran al golfo (Brenes & León, 1995; Brenes et al. 1996).
Durante el 2009, la profundidad promedio anual del disco de Secchi (3.36 ± 0.25 m, Cuadro 1) aumentó con respecto al 2008 y 2010. Esta profundidad es similar al máximo de 3.00 m encontrado por Brugnoli & Morales (1999) y Brugnoli-Olivera & Morales-Ramírez (2008) durante EN 1997.
En otros estudios en el Golfo de Nicoya, se observó que al aumentar la precipitación, la profundidad del disco de Secchi disminuye, mientras que la concentración de clorofila a aumenta (Calvo, 2002). Esta relación coincide con los resultados obtenidos en este estudio, pues durante el 2008 y el 2010 las precipitaciones promedios anuales fueron mayores, lo que resultó en menores profundidades del disco de Secchi y altas densidades de diatomeas y dinoflagelados; mientras que durante EN 2009, la precipitación promedio anual fue menor, aumentando así la profundidad del disco de Secchi y disminuyendo la densidad de diatomeas y dinoflagelados. De esta forma, el aumento en la profundidad del disco de Secchi en el 2009 (Fig. 4, Cuadro 1) probablemente se debió a la homogeneidad mensual de los sólidos totales durante el año, y a la baja densidad de los grupos microalgales provocada por una estabilidad ambiental por la disminución de la precipitación.
La precipitación promedio anual en el Golfo de Nicoya durante EN 2009 fue menor (79.86 ± 10.79 mm) que en LN 2008 y LN 2010 (Cuadro 1). Durante EN 1997-1998 (Punta Morales, Golfo de Nicoya) se reportó una precipitación acumulada de 1 357 mm, siendo el promedio anual de 113.08 mm (Brugnoli & Morales, 1999), aunque mundialmente EN 1997-1998 fue catalogado como uno de los más fuertes del siglo XX (Eissler & Quiñones, 1999; Lavaniegos et al. 2003; Palomares-García et al. 2003).
Las diatomeas y los dinoflagelados disminuyeron durante EN 2009 (Fig. 5A y B, Cuadros 2 y 3), a excepción de agosto y octubre de 2010, cuando los dinoflagelados superaron a las diatomeas a 1 y 5 m de profundidad. La mayoría de los autores han encontrado este comportamiento de las diatomeas con respecto a la presencia de EN en diferentes zonas marinas: Castillo & Vizcaíno (1993) en el Pacífico colombiano (EN 1982-1983), Blanchot et al. (1992) en el Pacífico ecuatorial (EN 1987), Lui et al. (1996) en el Pacífico ecuatorial (EN 1992), Rojas & Ortiz (2007) (EN 1998) en Colombia, y en la bahía Mejillones al sur de Chile (EN 1997-1998) (Eissler & Quiñones, 1999; Rodríguez, 2004). La disminución de la clorofila a y de la productividad primaria se han relacionado con EN (Escalante et al. 2013). Sin embargo, Lavaniegos et al. (2003) (EN 1997-1998) y Tapia (2007) (EN 1999-2000) observaron aumentos en las concentraciones de diatomeas. Con respecto a los dinoflagelados, la mayoría de los autores observaron una disminución en la densidad de ese grupo (Blanchot et al. 1992; Castillo & Vizcaíno, 1993; Torres-Zambrano, 1998; Rodríguez, 2004; Tapia, 2007), mientras que Lui et al. (1996) y Rojas & Ortiz (2007) reportaron más bien un aumento en su densidad. En el presente estudio, ocurrió una disminución en las densidades de los dinoflagelados, probablemente debido a la competencia o disponibilidad de los nutrientes.
En cuanto a las cianobacterias, durante EN 1987, Blanchot et al. (1992) determinaron una disminución en su densidad. En este estudio, concretamente a 1 m de profundidad, el promedio anual durante EN 2009 fue mayor que en LN 2008, y continuó aumentando en LN 2010, por lo que no se observó un comportamiento definido con respecto a la presencia de EN 2009. Esto probablemente ocurrió porque estas microalgas son capaces de competir ventajosamente con otros grupos en condiciones de ambientes extremos, lo que parece estar ligado a características de los ciclos del nitrógeno y del fósforo (Carmenate et al. 2010).
Quince especies de dinoflagelados disminuyeron sus concentraciones durante EN 2009. Algunas de ellas ya han sido catalogadas como bioindicadores de EN en diferentes regiones del Pacífico ecuatorial.
C. furca aparece como un dinoflagelado que aumenta sus concentraciones durante EN 1991-1992 (Castillo & Vizcaíno, 1993) y en EN 1997-1998 (Torres-Zambrano, 1998; Rojas & Ortiz, 2007). Durante EN 1997-1998, Brugnoli-Olivera & Morales-Ramírez (2008) únicamente mencionan al género Ceratium en el Golfo de Nicoya, sin describir especies ni la variación de sus concentraciones. Debido a su comportamiento en este estudio, esta especie debe tomarse en cuenta como bioindicadora potencial que disminuye o aumenta sus densidades durante los eventos cálidos de EN, en dependencia de la profundidad del Golfo de Nicoya.
C. fusus, al igual que C. furca, es un bioindicador que tiende a aumentar sus densidades durante los eventos de EN (Castillo & Vizcaíno, 1993; Rojas & Ortiz, 2007; Tapia, 2007), aunque en este estudio la disminución no fue significativa (P ˃ 0.05).
P. conicum disminuyó sus densidades en el Golfo de Nicoya durante el evento de EN 2009, y aunque fue reportado en el Pacífico suroeste peruano (Torres-Zambrano, 1998) y en el Pacífico colombiano (Rojas & Ortiz, 2007) durante el mismo evento EN 1997-1998, no se especificó su comportamiento durante el evento, al igual que Brugnoli-Olivera & Morales-Ramírez (2008) en el Golfo de Nicoya. C. tripos ha sido considerada una especie bioindicadora que aumenta sus densidades durante EN (Castillo & Vizcaíno, 1993; Torres-Zambrano, 1998; Rojas & Ortiz, 2007), al igual que G. polygramma (Torres-Zambrano, 1998; Rojas & Ortiz, 2007), pero al igual que las especies anteriores sus densidades disminuyeron en el Golfo de Nicoya durante EN 2009.
La diatomea S. costatum se ha considerado bioindicadora de los eventos EN, disminuyendo sus densidades en el Pacífico colombiano (Castillo & Vizcaíno, 1993) y aumentando en la costa peruana (Torres-Zambrano, 1998; Tarazona et al. 1999), durante EN 1997-1998. Brugnoli-Olivera & Morales-Ramírez (2008) midieron densidades altas en EN 1997, en la parte interna del Golfo de Nicoya. En este estudio, las densidades de S. costatum disminuyeron durante EN 2009.
Las especies anteriores han sido consideradas indicadoras de eventos EN. En el Golfo de Nicoya, estas especies disminuyeron sus densidades durante EN 2009 y aumentaron durante LN 2008 y LN 2010. Otras especies que disminuyen su densidad, pero que no han sido descritas como indicadoras de EN fueron: G. catenatum, A. fraterculus, A. monilatum, D. caudata, N. scintillans, Tipo 1, C. lineatum, G. verior y L. polyedrum.
P. micans aumentó sus densidades durante EN 2009 en el Golfo de Nicoya, y ha sido considerada bioindicadora por Rodríguez (2004), al exhibir altas densidades en la bahía Mejillones (Antofagasta, Chile, EN 1997-1998), al igual que en el Pacífico colombiano (Rojas & Ortiz, 2007) y peruano (Torres-Zambrano, 1998). Los resultados encontrados en el presente estudio durante EN 2009 concuerdan con lo observado por otros autores en diferentes regiones del Pacífico ecuatorial. Los dinoflagelados C. macroceros, P. kofoidii, y las cianobacterias T. erythraeum y M. elegans, aumentaron sus densidades durante EN 2009 en el Golfo de Nicoya, sin embargo, no han sido reportadas hasta la fecha como bioindicadoras de EN en otras regiones.
De las especies exclusivas durante el evento EN 2009 en el Golfo de Nicoya, solo P. bipes (Castillo & Vizcaíno, 1993; Torres-Zambrano, 1998; Rodríguez, 2004) y C. massiliense (Castillo & Vizcaíno, 1993; Rojas & Ortiz, 2007) han sido descritas en diferentes eventos EN (1991-1992 y 1997-1998). Sin embargo, otras especies que podrían ser consideradas bioindicadoras exclusivas de eventos EN son los dinoflagelados G. instriatum, Ceratocorys sp. y P. balticum, y la diatomea Aulacodiscus sp.
Por último, algunas especies estuvieron ausentes durante EN 2009 en el Golfo de Nicoya (Cuadros 4 y 5), no así en otras regiones. C. trichoceros estuvo presente y aumentó su densidad en el Pacífico colombiano durante EN 1991-1992 (Castillo & Vizcaíno, 1993), mientras que durante EN 1997-1998, en la bahía Mejillones (Chile), disminuyó su densidad (Rodríguez, 2004). C. macroceros disminuyó su densidad en el Pacífico colombiano durante EN 1991-1992 (Castillo & Vizcaíno, 1993). C. lineatum, O. steinii y Alexandrium sp. estuvieron ausentes durante EN 2009, este comportamiento les podría atribuir un rango de bioindicadoras potenciales en el Golfo de Nicoya.
CONCLUSIONES
En general, durante el evento atmosférico EN 2009, las propiedades fisicoquímicas variaron significativamente con respecto a las registradas durante los eventos LN 2008 y 2010.
El evento EN 2009 causó variaciones importantes en la comunidad microfitoplanctónica del Golfo de Nicoya. Entre ellas se destacan: (1) la disminución de la densidad de los dinoflagelados: G. catenatum, A. fraterculus, A. monilatum, D. caudata, N. scintillans, C. furca, Tipo 1, C. fusus, P. conicum, G. verior, C. tripos, L. polyedrum, C. lineatum, G. polygramma, y la diatomea S. costatum; (2) aumento de la densidad de los dinoflagelados: C. macroceros, P. kofoidii, P. micans, C. furca, y las cianobacterias: T. erythraeum y M. elegans; (3) presencia exclusiva de dinoflagelados: G. instriatum, Ceratocorys sp., P. bipes, C. massiliense, G. instriatum, P. balticum, y la diatomea Aulacodiscus sp. (4) ausencia de C. lineatum, C. trichoceros, O. steinii, Alexamdrium sp. y C. macroceros.
Esas variaciones en las concentraciones y asociaciones de especies de microalgas durante LN 2008, EN 2009 y LN 2010, pueden ser utilizadas como potenciales indicadores biológicos marinos de los eventos atmosféricos de EN y LN.
AGRADECIMIENTOS
Los autores agradecen a la Universidad Nacional el apoyo para la realización de este estudio. Parte de este trabajo fue financiado con fondos de la Ley de Pesca y Acuicultura No. 8436. Al Instituto Meteorológico Nacional de Costa Rica por el aporte de los datos de precipitación y épocas de EN y LN. A Claudio Calvo Vargas, por sus observaciones y aportes tan valiosos. Al Capitán Orlando Torres Castro por la colaboración en la recolecta de las muestras. Deseamos agradecer la revisión realizada por los revisores anónimos al manuscrito, lo que contribuyó al enriquecimiento del mismo.

Efectos de El Niño y La Niña sobre el comportamiento del microfitoplancton marino y las variables fisicoquímicas durante el 2008 a 2010 en el Golfo de Nicoya, Costa Rica por Revista Ciencias Marinas y Costeras se distribuye bajo una Creative Commons Reconocimiento-NoComercial-SinObraDerivada 3.0 Costa Rica License.
Basada en una obra en http://www.revistas.una.ac.cr/index.php/revmar.
Permisos que vayan más allá de lo cubierto por esta licencia pueden encontrarse en revmar@una.cr.
BIBLIOGRAFÍA
Aguirre-Gómez, R., Salmerón, O. & Álvarez, R. (2003). Effects of ENSO off the southwest coast of México, 1996-1999. Geofís. Int., 42(3), 377-388.
Badan, A. (2003). The effects of El Niño in Mexico: A survey. Geofís. Int., 42(3), 567-571.
Blanchot, J., Rodier, M. & Le Bouteiller, A. (1992). Effect of El Niño Southern Oscillation events on the distribution and abundance of phytoplankton in the Western Pacific Tropical Ocean along 165°E. J. Plankton Res., 14(1), 137-156. doi: http://dx.doi.org/10.1093/plankt/14.1.137
Boening, C., Willis, J. K., Landerer, F. W. & Nerem, R. S. (2012). The 2011 La Niña: So strong, the oceans fell. Geophys. Res. Lett., 39, 1-5. doi:10.1029/2012GL053055
Brenes, C. & León, S. (1995, noviembre). Hidrografía del Golfo de Nicoya, Costa Rica. Ponencia presentada en el Simposium Ecosistema de manglares en el pacífico centroamericano, San Salvador, El Salvador.
Brenes, C., León, S. & Arroyo, G. (1996). Influence of coastal waters on some physical and chemical oceanographic characteristics of Gulf of Nicoya, Costa Rica. Top. Meteor. Oceanogr., 3(2), 65-72.
Brugnoli, O. E. & Morales, R. A. (1999). Parámetros físico-químicos de la columna de agua, en la zona de Punta Morales, Golfo de Nicoya, durante un evento “El Niño”. Top. Meteor. Oceanog., 6(2), 52-57.
Brugnoli-Olivera, E. & Morales-Ramírez, A. (2008). Trophic planktonic dynamics in a tropical estuary, Gulf of Nicoya, Pacific coast of Costa Rica during El Niño 1997 event. Rev. Biol. Mar. Oceanogr., 43(1), 75-89. doi: http://dx.doi.org/10.4067/S0718-19572008000100008
Calvo, E. (2002). Factores bióticos y abióticos relacionados con la distribución del dinoflagelado tóxico Pyrodinium bahamense var. compressum (Plate 1906) en el Golfo de Nicoya, Costa Rica. Tesis de licenciatura no publicada. Universidad Nacional, Heredia, Costa Rica.
Carmenate, M., Arriaza, L., Busutil, L., Durán, A., García, C. & García, I. (2010). Calidad del agua marina en un tramo costero con uso industrial de la provincia La Habana. Recuperado en marzo 29, 2012, disponible en http://www.oceandocs.org/bitstream/1834/
Castillo, F. A. & Vizcaíno, B. Z. (1993). Observación del fitoplancton del Pacífico colombiano durante 1991-1992 en condiciones El Niño. Bull. Inst. Fr. études andines, 22(1), 179-190.
Castro, E. & León, S. (1998). El estado del arte de la investigación en valoración de aguas degradadas en regiones marino-costeras: El caso del Golfo de Nicoya. Informe Preliminar. Heredia, Costa Rica. Centro Internacional de Política Económica para el Desarrollo Sostenible-CINPE.
Eissler, Y. & Quiñones, R. A. (1999). Microplanktonic respiration off northern Chile during El Niño 1997-1998. J. Plankton Res., 21(12), 2263-2283. doi: http://dx.doi.org/10.4067/S0718-19572008000100008
Escalante, F., Valdez-Holguin, J. E., Álvarez-Borrego, S. & Lara-Lara, J. R. (2013). Temporal and spatial variation of sea surface temperature, chlorophyll a, and primary productivity in the Gulf of California. Ciencias Marinas, 39(2), 203-215. doi: http://dx.doi.org/10.7773/cm.v39i2.2233
Filonov, A. E., Tereshchenko, I. E. & Monzón, C. O. (2003). Hydrographic monitoring of El Niño 97-98 off the coast of southwest Mexico. Geofís. Int., 42(3), 307-312.
Gajardo, N., González, H. E. & Marchant, M. (2013). Characterization of El Niño, La Niña, and normal conditions through planktonic foraminifera (2006-2007) in the southeastern Pacific. Ciencias Marinas, 39(3), 253-264. doi: http://dx.doi.org/10.7773/cm.v39i3.2177
García, D. X. F., Gusmão, L. M. de O. & Herrera, Y. (2008). Influencia de los eventos climáticos El Niño y La Niña en la comunidad de Chaetognatha de las aguas superficiales del océano Pacífico colombiano. Rev. Bras. Enga. Pesca, 3(1), 30-50.
Gaxiola-Castro, G., Durazno, R., Lavaniegos, B., De la Cruz-Orozco, M. E., Millán-Núñez, E., Soto-Mardones, L. & Cepeda-Morales, J. (2008). Pelagic ecosystem response to interanual variability off Baja California. Ciencias Marinas, 34(2), 263-270.
Lavaniegos, B. E., Gaxiola-Castro, G., Jiménez-Pérez, L. C., González-Esparza, M. R., Baumgartner, T. & García-Córdova, J. (2003). 1997-98 El Niño effects on the pelagic ecosystem of the California Current off Baja California, Mexico. Geofís. Int., 42(3), 483-494.
León-Coto, S., Kress, N., Brenes-Rodríguez, C. L. & Brenner, S. (1998). Una contribución a la ecología del Golfo de Nicoya. Uniciencia, 15(16), 35-37.
Lui, D., Fryxell, G. A. & Kacsmarska, I. (1996). El Niño (1992) in the equatorial Pacific: low biomass with a few dominating species in the microphytoplankton. J. Plankton Res., 18(7), 1167-1184. doi: http://dx.doi.org/10.1093/plankt/18.7.1167
Magaña, V. O., Vázquez, J. L., Pérez, J. L. & Pérez, J. B. (2003). Impact of El Niño on precipitation in México. Geof. Int., 42(3), 313-330.
Morales-Ramírez, A. & Brugnoli-Olivera, E. (2001). El Niño 1997-1998 impact on the plankton dynamics in the Gulf of Nicoya, Pacific coast of Costa Rica. Rev. Biol. Trop., 49(2), 103-114.
NOAA. (2011). El Niño and La Niña years and Intensities. Based on Oceanic Niño Index (ONI). Recuperado en diciembre 5, 2011, disponible en http://www.cpc. noaa.gov/products/analysis_monitoring/ensostuff/ensoyears.shtml
Ochoa, J. L. (2003). ENSO phenomenon and toxic red tides in México. Geofís. Int., 42(3), 505-515.
Palomares-García, R., Martínez-López, A., De Silva-Dávila, R., Funes-Rodríguez, R., Carballido-Carranza, M. A., Avendaño-Ibarra, R., Hinojosa-Medina, A. & López-Ibarra, G. A. (2003). Biological effects of El Niño 1997-98 on a shallow subtropical ecosystem: Bahía Magdalena, México. Geofís. Int., 42(3), 455-466.
Reguera, B. (2002). Establecimiento de un programa de seguimiento de microalgas tóxicas. En E. A. Sar, M. E. Ferrario & B. Reguera (Eds.), Floraciones algales nocivas en el Cono Sur Americano (pp. 21-53). Madrid, España: Instituto Español de Oceanografía.
Rodríguez, L. (2004). Observaciones sobre efectos de El Niño 1997-1998 en el fitoplancton de bahía Antofagasta y durante un fenómeno de surgencia en bahía Mejillones del Sur, Antofagasta, Chile. En S. Avaria, J. Carrasco, J. Rutllant & E. Yáñez (Eds.), El Niño - La Niña 1997-2000. Sus efectos en Chile (pp. 97-118). Valparaíso, Chile: CONA.
Rojas, H. P. J. & Ortiz, J. R. (2007). Comportamiento del fitoplancton durante el evento ENOS en el Océano Pacífico colombiano. Ing. Rec. Nat. Amb., 6, 5-15.
SPSS. (2006). Statistical Package for the social science for Windows, version 17.0. Chicago, EE. UU.: SPSS Inc.
Tapia, M. E. (2007). Variabilidad temporal del fitoplancton en áreas costeras del mar ecuatoriano y su interrelación con el evento La Niña 1999-2000, Ecuador. Acta Oceanogr. Pacífic., 14(1), 37-48.
Tarazona, J., Indacochea, A., Valle, S., Córdova, C., Ochoa, N., Serrano, W. & Peña, T. (1999). Impacto de “El Niño 1997-98” sobre el ecosistema marino somero de la costa central del Perú. Rev. Peru. Biol., Vol. extraordinario, 18-31.
Todd, R. E., Rudnick, D. L., Davis, R. E. & Ohman, M. D. (2011). Underwater gliders reveal rapid arrival of El Niño effects off California’s coast. Geophys. Res. Lett., 38, 1-5. doi:10.1029/2010GL046376
Torres-Zambrano, G. (1998). Distribución cuali-cuantitativa del fitoplancton en el Pacífico sureste (3°16’-31°23’ S), durante El Niño 1997-1998. Acta Oceanogr. Pacífic., 9(1), 95-102.